
Abstract
Asexual reproduction by fissiparity has only been described from very few of the approximately 1300 nemertean species that otherwise mainly reproduce sexually. The best studied fissiparous species, Lineus sanguineus (Rathke 1799), is a cosmopolitan heteronemertean species inhabiting intertidal habitats of temperate coasts. Although sexual reproduction has never been described, molecular data suggest that sexual reproduction substantially shapes the genetic structure of the investigated populations. In an attempt to clarify the extent of sexual reproduction, three gene fragments (COI, 16S, ITS) were sequenced for 108 specimens sampled in 8 European localities. The results of a phylogenetic analysis and haplotype network showed no clear distinction between different populations, thus indicating the presence of sexual reproduction. Furthermore, we provide circumstantial evidence for the presence of a comparably long-lived planktonic larval stage as present in the closest relatives of L. sanguineus. To further understand the impact of abiotic factors on sexual reproduction and fissiparity, the effect of different temperature and illumination regimes on reproductive behavior and fragmentation was studied in specimens from a population from Bergen, Norway that share the same haplotype. Experimental setups represented summer (long light period and elevated temperature) and winter (short light and decreased temperature) conditions. Under winter conditions, a higher number of animals remained sexually mature and at least one specimen shed eggs on one occasion. Thus, although short light and/or low temperatures are most likely the influential factors on sexual maturity, the factors that influence fissiparity are less clear. The results of this study further solidify the cosmopolitan status of L. sanguineus and clarify the population structuring of this species. In addition, the study provides first data on the dynamics of sexual and asexual reproduction modes on which future investigations will have to expand, especially regarding genetic and physiological aspects.
Similar content being viewed by others


Introduction
In the animal kingdom, many species are capable of regenerating lost body parts, with regeneration of posterior body parts being fairly common (Bely and Nyberg 2010). On the other hand, fewer lineages are capable of regenerating a head. Within Nemertea, several species have independently gained this remarkable ability, one of the latest examples being Baseodiscus delineatus examined in Japan where male specimens reproduce by fissiparity in populations that appear to lack females (Ikenaga et al. 2019; Zattara et al. 2019). The best studied nemertean species in this respect is Lineus (= Ramphogordius) sanguineus that has already been studied for more than 200 years for its spontaneous fragmentation and regenerative capacities (e.g. McIntosh 1873–1874; Coe 1929, 1930b; Sivaradjam and Bierne 1981; Bierne 1990). Individuals of this species are known to naturally fragment into small pieces under certain conditions. These include warm temperatures, abundant food, and enough oxygen (Reutter 1967). From each fragment containing parts of the central nervous system, a complete individual can be regenerated, including the regeneration of a new head (Coe 1929, 1930b, 1934). These outstanding regenerative abilities are thought to provide the basis for asexual reproduction by fissiparity, which appears to be the dominant mode of reproduction in this species (Gontcharoff 1951; Bierne 1970; Gibson 1972).
Lineus sanguineus is a cosmopolitan heteronemertean species occurring in temperate waters (Riser 1994; Gibson 1995; Caplins and Turbeville 2011; Runnels 2013; Kang et al. 2015). Distribution records include European coasts, Atlantic and Pacific coasts of North and South America, and north Asian Pacific coasts (Gibson 1982, 1995; Kang et al. 2015). The species is found in the intertidal zone under rocks and stones, and in association with mussels and oysters on rocky shores (McIntosh 1873–1874; Coe 1943; Gibson 1982, 1995; Caplins and Turbeville 2011; Hiebert and Hunt 2015). Lineus sanguineus presents strong intraspecific color variation rendering species identification difficult (e.g. McIntosh 1873–1874; Gontcharoff 1951; Riser 1993, 1994; Caplins and Turbeville 2011). Due to its external appearance, L. sanguineus is often confused with the similar-looking species Lineus ruber, Lineus viridis or Lineus clandestinus (Kang et al. 2015; Krämer et al. 2017). In most phylogenetic analyses, L. sanguineus formed part of the “L. ruber” complex (Sundberg and Saur 1998; Thollesson and Norenburg 2003; Andrade et al. 2012), but more recent molecular analyses revealed a closer relationship to Riseriellus occultus, Lineus lacteus and Lineus longissimus (Ament-Velásquez et al. 2016). The closest related sister group is Lineus (= Ramphogordius) pseudolacteus, a hybrid species of Lineus lacteus and Lineus sanguineus that has so far only been recorded in Brittany (Gontcharoff 1951; Ament-Velásquez et al. 2016).
Due to the cosmopolitan distribution of L. sanguineus, the species was formerly seen as four separate species based on their respective distribution: L. sanguineus (European Atlantic coasts), Lineus nigricans (Mediterranean Sea), Lineus socialis (North American Atlantic coasts), and Lineus vegetus (North American Pacific coasts) (Gibson 1995). More recently, several morphological and molecular investigations concluded that there is only one species with a wide distributional range (Bierne et al. 1993; Riser 1993; Runnels 2013; Kang et al. 2015). This contradicts a generally observed phenomenon whereby assumed cosmopolitan species hide cryptic speciation, thus significantly reducing the distributional range of each of the species (e.g. Dawson and Jacobs 2001 in cnidarians, Gómez et al. 2002 in rotifers, and Leasi et al. 2016 in the hoplonemertean genus Ototyphlonemertes). This unusual cosmopolitan distribution can either be the result of long-distance dispersal via long-lived planktonic larvae or by dispersal of adult specimens or encysted fragments (Riser 1993, 1994; Caplins and Turbeville 2011; Kang et al. 2015). Adults could be transported by natural (rafting) or by anthropogenic means (in the fouling community of ship hulls) (Runnels 2013; Kang et al. 2015). Due to the so-far lacking evidence of a planktonic larvae, dispersal by ship was suggested as the most likely dispersal vector for a predominantly asexually reproducing species, such as L. sanguineus (Riser 1994; Runnels 2013; Kang et al. 2015).
This asexual reproduction by fissiparity sets L. sanguineus apart from other closely related species as neither species, except for L. pseudolacteus, is known to reproduce asexually. A reproductive system that is solely based on asexual reproduction also has a severe impact on population structuring as no larvae are present that could account for long-distance dispersal and genetic exchange ensuring little genetic structuring between remote populations (Jablonski 1986; Palumbi et al. 1997; Avise 2000; Cahill et al. 2017). One therefore would expect significant genetic structuring between distant populations, but only low levels of genetic diversity within the populations if the species reproduced solely asexually (Johnson and Threlfall 1987; Palumbi 1992; Bürger 1999). So far only few studies of population structure on L. sanguineus that could give evidence about the amount of asexual reproduction exist although it is a cosmopolitan species and has been studied extensively for more than 200 years (Kang et al. 2015; Ament-Velásquez et al. 2016; Zattara et al. 2019).
Based on current genetic evidence, sexual reproduction has to take place in L. sanguineus at least to some extent, although there is no definite proof of it (Riser 1994; Kang et al. 2015; Ament-Velásquez et al. 2016). In this study, we sequenced gene fragments of the mitochondrial cytochrome c oxidase subunit I (COI) and 16S rRNA (16S) as well as the nuclear internal transcribed spacer region (ITS) to examine population structure and haplotype diversity. Moreover, we performed phylogenetic analyses based on each gene fragment and a combined dataset. The results were analyzed with special emphasis on indication of sexual reproduction.
All of the more closely related heteronemertean species possess long-lived, planktonic pilidium larvae (e.g. Beckers et al. 2015 for R. occultus). This leads to the assumption that some kind of larvae has to be present in L. sanguineus, too. As no mature specimens were ever found between May and October, sexual reproduction was assumed to take place between mid-winter and early spring in the northern hemisphere (McIntosh 1873–1874; Coe 1899; Gontcharoff 1951; Bierne 1970; Gibson 1972; Riser 1994). Spermatogenesis seems to take place in a regular manner (Gontcharoff 1951; Döhren et al. 2010), whereas oogenesis is often reported as abortive, especially in specimens sampled in Brittany (Coe 1943; Gontcharoff 1951; Riser 1994). Thus, there are no observations of fertilization or egg deposition, let alone larval development of L. sanguineus. Observations on the closely related species L. lacteus and L. ruber revealed that photoperiod or temperature may have an effect on gonad maturation, with low temperatures and long periods of darkness triggering gonadogenesis (Vernet and Bierne 1988, 1993). Based on these findings we decided to further investigate the effect of light and temperature on sexual maturation in specimens from a Northern-European population of L. sanguineus sharing the same haplotype expecting low temperatures and a short photoperiod to promote sexual maturity. Twenty specimens produced by asexual reproduction were used. Fragmentation, growth and the state of sexual maturity were recorded for several months under different temperature and light regimes.
Material and methods
Specimens and sampling sites
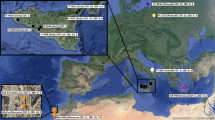
108 specimens of Lineus sanguineus (Rathke, 1799) were collected between 2010 and 2020 from eight European localities (Fig. 1). These included Germany (Helgoland, Sylt), Norway (Bergen), and France (Banyuls, Concarneau, Ile de Groix, Roscoff, Wimereux). Specimens were found either under rocks, in rock fissures or associated with mussels (Mytilus edulis). Photographs were taken with a digital camera (Canon EOS 600D) mounted on a dissection microscope (Zeiss Stemi 2000). A tissue sample of each individual was preserved in absolute ethanol (99.9%) for DNA extraction. Specimens sampled in 2014 in Bergen were collected alive and kept at the Institute of Evolutionary Biology in Bonn at 18 °C and under a light-darkness regime of 16 h:8 h for the experiments on reproduction.

Specimens of Lineus sanguineus were collected from eight European localities, including five localities in France, two localities in Germany, and one locality in Norway
Nucleic acid purification and PCR amplification
DNA extraction was performed using the DNeasy Blood and Tissue Kit (Qiagen) following the manufacturer’s extraction protocol. For this study, three loci were amplified: the partial mitochondrial COI and 16S rRNA and the nuclear ITS gene fragment (ITS1, 5.8S rRNA, and ITS2) using the following primers: for COI LCO1490 [GGTCAACAAATCATAAAGATATTGG] and HCO2198 [TAAACTTCAGGGTGACCAAAAAATCA] (Folmer et al. 1994), for 16S 16Sar-L [CGCCTGTTTATCAAAAACAT] and 16Sbr-H [CCGGTCTGAACTCAGATCACGT] (Palumbi et al. 1991), and for ITS ITS-28S [TTTTCAACTTTCCCTCACGG] and ITS-18S [CATTTGAGGAAGTAAAAGTCGTAAC] (Krämer et al. 2017). Polymerase chain reactions for COI and 16S were performed using Hot-Master Taq polymerase (Invitrogen™) whereas Dream Taq™ PCR Master Mix (Thermo Fisher) was used for ITS. PCR cycling was initiated with 2 min at 94 °C, followed by 40 cycles (30 s at 94 °C, 60 s at 48 °C and 60 s (COI, 16S)/120 s (ITS) at 72 °C), and terminated with a 2 min (COI, 16S)/ 1 min (ITS) final elongation at 72 °C. Amplified products were purified using illustra ExoProStar 1-Step (GE Healthcare) following the manufacturer’s instructions. Sanger sequencing was conducted by LGC Genomics (Berlin, Germany) using only forward primers (COI, 16S) and forward and reverse primers (ITS) for sequencing (Sanger et al. 1977). All sequences have been deposited in the GenBank database (accession numbers provided in Table 1).
Sequence analysis
Sequences were edited with BioEdit version 7.2.5 (Hall 1999) and aligned using MAFFT version 7 (Katoh et al. 2019). For sequence alignment G-INS-I strategy with default parameters (scoring matrix for nucleotide sequences: 200PAM/K = 2, gap opening penalty: 1.53, offset value: 0.0) were chosen. All ambiguous positions were excluded with Gblocks version 0.91b (Castresana 2000) using default parameters.
For phylogenetic analyses based on the combined COI (512 nt), 16S (398 nt), and ITS (967 nt) data, 59 Lineus sanguineus specimens were selected to represent all European sampling sites except for Banyuls. Furthermore, COI and 16S data were combined with sequence data taken from GenBank (for Accession numbers see Supplementary Table 1), whereas ITS data was analyzed separately. For all four approaches the model for phylogenetic reconstruction was selected by MrModeltest2 version 2.3 based on the Akaike information criterion (Nylander 2004). General-time-reversible model and a gamma distribution with a proportion of invariant sites (GTR + G + I) were selected as best-fitting substitution model for the COI and 16S dataset (Nei and Kumar 2000), whereas Hasegawa-Kishino-Yano model and a gamma distribution of invariant sited (HKY + G + I) were selected for the ITS and the combined dataset (Hasegawa et al. 1985). Phylogenetic trees were reconstructed in MEGA version 6.06 using a maximum likelihood (ML) as optimality criterion (Tamura et al. 2013). Branch support was estimated using 500 bootstrap replicates. Lineus ruber, Lineus viridis, and Lineus clandestinus were used as outgroups.
Haplotype networks for the COI, 16S, and ITS datasets were estimated using statistical parsimony implemented in TCS version 1.21 (Clement et al. 2000) with the connection limit set to 95% (Templeton et al. 1992). Haplotype and nucleotide diversity were calculated using DnaSP version 6.12.03 (Rozas et al. 2017). Analysis of Molecular Variance (AMOVA) was utilized to examine genetic variation within and among populations (Excoffier et al. 1992). The analyses were performed with GenAlEx version 6.503 (Peakall and Smouse 2006, 2012), using 999 permutations to test the significance. A Mantel test was also performed using GenAlEx to determine the correlation between genetic and geographic distances of populations (Mantel 1967).
Fragmentation/sexual maturity experiments
Specimens of L. sanguineus from Bergen (Norway) were collected in 2014. Until the start of the experiments in 2018, the worms were kept at constant temperature (18 °C) and 16 h of light in plastic boxes with approx. one liter of standing sea water. The water was changed every other week. The animals were fed with Tubifex tubifex (Müller, 1774) once a week. To gain information on the influence of temperature and light on fragmentation and sexual maturation, two experimental setups were designed: two sets of 20 worms each, were kept together in plastic boxes with 0.5 L of aerated artificial sea water under the contrasting influence of winter conditions (9 °C, 8 h light) and summer conditions (18 °C, 16 h light). All specimens had the same haplotypes of the markers used herein as identified by sequence analysis. During the 6-month duration of the experiment, the worms were fed once a week and the water was changed once every week. At the onset of the experiments and afterward each month, the complete worms, defined as possessing a cephalic lobe (i.e. head) and the fragments were counted and measured, and the sexual development was recorded. For these procedures, the animals were immobilized by immersion in 3,5% MgCl2 in seawater (mixed from equal volumes of 7% MgCl2-solution and artificial sea water) for a minimum of 15 min. That causes the entire musculature to relax and thus allows for standardized measurements. Specimens were regarded as sexually mature when gonads and gonopores were well visible. Detailed results of the counting and measurement are provided in Supplementary Tables 2 and 3.
The results were tested for normal distribution in R version 3.6.1 (R Core Team 2019). None of the datasets were normally distributed. Non-parametric Mann–Whitney U test was performed to compare animal length under the different conditions. Results were regarded as significant when p < 0.05. Boxplots were computed using the ggplot2 package (Wickham 2016).
Microscopy of larvae
In April 2013, larvae were found during a water change in the container with specimens collected in Roscoff, France kept at 18 °C with 16 h of light. Due to the small number and size of the larvae, they were not fixed for DNA extraction, but documented with light microscopic methods. After immobilization of the musculature by immersion in 3,5% MgCl2 in seawater, the larvae were mounted in the same mixture on a glass slide. To prevent them from swimming around with ciliary action, the larvae were slightly compressed with the cover slip mounted on clay feet. The mounted larvae were examined with a BX 51 everse microscope (Olympus) using differential interference contrast with a 40 × , NA 0.75 dry objective. Images were taken with a ColorView IIIu CCD camera (Soft Imaging System). Images were further processed and mounted with Adobe Photoshop CS5 and Illustrator CS5 (Adobe).
Results
Phylogenetic analysis
No resolution of different geographic Lineus sanguineus populations was visible in the ML analyses of any of the single genes or the combined gene set. All included specimens form one well-supported clade. Because of this, we only show the ML analysis based on the combined COI, 16S, and ITS sequences (Fig. 2). The Norwegian population from Bergen (represented by one specimen) that has been used in the fragmentation/sexual maturity experiments forms one clade with three specimens from Sylt and three specimens from Roscoff. Besides this, specimens from all localities are mixed.

Maximum likelihood tree (HKY + G + I) of selected Lineus sanguineus specimens based on concatenated COI mtDNA, 16S rRNA, and ITS rRNA sequences. Lineus ruber, Lineus viridis, and Lineus clandestinus were used for outgroup rooting. Numbers above nodes indicate bootstrap support from 500 replicates for each clade. Only values above 90 are shown. Specimens from both further examined populations are highlighted (blue = Bergen, Norway)
Population analysis
We obtained COI sequences for 104 specimens from eight European localities which resulted in an alignment of 512nt in length. These sequences were combined with 183 Lineus sanguineus specimens deposited at GenBank. These GenBank specimens originated from different localities in Europe, the Pacific coast of Canada, China, Argentina, and Chile. The whole dataset comprised 15 polymorphic sites. Haplotype diversity was moderately high (Hd = 0.732; SD = 0.011), but nucleotide diversity was very low (π = 0.00315; SD = 0.00016).
The haplotype network based on the COI dataset yielded 15 haplotypes (Fig. 3A). Approximately 90% of all individuals possess one of three highly frequent haplotypes (72, 86, and 89 specimens, respectively). Specimens with these haplotypes originate from all sites sampled for this study, as well as from other locations in Europe, Pacific Canada, China, Argentina, and Chile. There is no separation of haplotypes by locality although all specimens from the Mediterranean share the same haplotype (H4). The specimens used in the fragmentation/sexual maturity-experiments from Bergen possess haplotype H1.

taken from GenBank. Geographic distribution by maritime zones is represented by color. Specimens from Bergen, Norway used in fragmentation/sexual maturity-experiments are highlighted (blue). Geographic distribution in C is based on the sampling sites. Numbers within pie charts represent number of specimens with respective haplotype. Empty lines indicate a single substitution; each black dot indicates one additional mutational step
Statistical parsimony haplotype networks of all sequenced Lineus sanguineus specimens with a connection limit of 95% based on mitochondrial cytochrome c oxidase subunit I gene (A), mitochondrial 16S rRNA (B), nuclear ITS rRNA (C), and a concatenated dataset including all three gene fragments (D). A and B include additional sequence data
The analysis of the 398-nt-long alignment of 16S was based on 86 specimens sampled for this study and 16 sequences taken from GenBank. The data deposited at GenBank originates from individuals sampled in Buenos Aires, Oregon, and Wales. In this dataset, 11 polymorphic sites were present. Haplotype diversity was moderate (Hd = 0.535; SD = 0.037), but nucleotide diversity was very low (π = 0.00204; SD = 0.00037).
The haplotype network based on all 105 specimens comprised seven haplotypes (Fig. 3B). Approximately 80% of all individuals have one of the two highly frequent haplotypes (H1: 57 specimens, H2: 30 specimens; the latter also found in specimens from Bergen used in the fragmentation/sexual maturity-experiments). At least some specimens of each population have one of the two most frequent haplotypes. The remaining seven haplotypes occur in only one to four individuals, respectively. As in the network obtained from COI data, haplotypes are not clustered based on locality.
We obtained ITS sequences for 60 specimens from seven European localities that resulted in an alignment of 967nt in length. In total, 26 polymorphic sites were present. Haplotype diversity was high (Hd = 0.885; SD = 0.024), whereas nucleotide diversity was low (π = 0.00527; SD = 0.00032).
The statistical parsimony analysis of the ITS dataset yielded one haplotype network represented by 17 haplotypes (Fig. 3C). There are no highly frequent haplotypes, but seven medium frequency haplotypes found in three to six individuals, respectively. Six of these are shared by specimens from at least two different locations. The remaining ten haplotypes are represented by one specimen each. Interconnections to the closest haplotype vary between one and six substitutions. In comparison to the haplotype networks generated by the COI and 16S datasets, haplotype diversity and structuring is most pronounced in the ITS dataset.
A fourth statistical parsimony analysis was based on a concatenated dataset of all three genetic markers comprising 58 specimens from seven European localities. The analysis yielded one network represented by 22 haplotypes (Fig. 3D). The two most frequent haplotypes (H1: 14 specimens, H2: 9 specimens) includes specimens of the further investigated population from Bergen. Both are separated by 25 nucleotide substitutions. Four of the five medium-frequency haplotypes found in three, four and five specimens, respectively, are shared by specimens from at least two different locations. The fifth haplotype (H5) is only found in 4 specimens collected in Helgoland. The remaining 15 haplotypes are only found in one specimen each. As for the ITS dataset, the combined dataset also shows more pronounced haplotype structuring compared to COI and 16S.
Based on the AMOVA analysis of the COI dataset there is a higher level of differentiation among populations (ΦPT = 0.327; p < 0.05) than among regions (ΦRT = 0.109; p < 0.05). This result hints at gene flow among the regions, although structuring is to some extent present among populations. Only 11% of all variation is found among regions, 29% is found among populations, and 60% within populations indicating that variation between individuals is higher than variation between populations. Pairwise ΦPT values were obtained by combining close geographical populations into 8 regions (Table 2). The three different northern Atlantic regions showed low levels of differentiation from each other indicating a high amount of gene flow between these populations (ΦPT = 0.016–0.085, p < 0.05). Comparably low levels of differentiation are inferred between Chile and Pacific Canada, Chile and China, and Argentina and the north eastern Atlantic region (ΦPT = 0.000–0.034, p > 0.05) although these results are not significant due to the small number of specimens present for Chile and Argentina. Low to moderate differentiation is present between Chinese and northern Atlantic and Canadian Pacific populations (ΦPT = 0.03–0.153, p < 0.05). Moderate differentiation was inferred between Canadian Pacific populations and the three northern Atlantic regions (ΦPT = 0.230–0.422, p < 0.05). Significantly high levels of differentiation can be observed between populations from Norway and north western Pacific coasts, and between Chilean and northern Atlantic populations (ΦPT = 0.415–0.98; p < 0.05). Populations from the Mediterranean on the other hand are highly differentiated from all other regions indicating reduced gene flow between the Mediterranean and the other regions (ΦPT = 0.618–0.785, p < 0.05). This high level of differentiation between the Mediterranean and all other population might account for the relatively high differentiation among all included populations (ΦPT = 0.327; p < 0.05). In the Mantel test, there was no significant relationship between geographic and genetic distance (Rxy = 0.047, p > 0.05), thus excluding isolation by distance.
Fragmentation
In the Bergen population, fragmentation starts earlier under winter conditions and also yields more fragments than under summer conditions (Fig. 4A). The number of specimens that regenerated a head under winter conditions does not significantly increase. Summer conditions, on the other hand, delay fragmentation until month 3 and the total amount of generated fragments is always lower than under winter conditions (Fig. 4A). Nevertheless, the total amount of individuals that successfully regenerated a head is significantly higher under summer conditions (Month 6: 32 summer vs. 20 winter). At the beginning of the experiments all specimens used for the different setups where approximately the same size (Mann–Whitney U test, W = 160.5, p = 0.3). At the end of experiments specimens kept at winter conditions are significantly longer than specimens kept at summer conditions (Mann–Whitney U test, W = 182, p = 0.007). Growing in length commences later at winter conditions than at summer conditions but at summer condition there is an overall strong decrease in length over time with the majority of specimens ranging below 50 mm in length after three months (Fig. 4B).

Fragmentation and growth of Lineus sanguineus specimens from the Bergen, Norway population at summer (18 °C, 16 h light) and winter conditions (9 °C, 8 h light). Experimental procedures commenced with 20 individuals for each of the two setups. Complete animals and fragments were counted and measured for six months. A Amount of fragments and specimens with head of L. sanguineus. B Length variation of specimens with head over a period of 6 months. Significance was tested based on the Wilcoxon rank sum test (*p < 0.05, **p < 0.01)
Sexual maturation and spawning
In the specimens from Bergen, gonads were already present in approximately 2/3 of all specimens at the start of the experiment (Fig. 5A). During the course of the experiment though, the number of specimens bearing gonads at summer conditions drastically declined. In contrast to this, nearly half of the specimens at winter conditions had gonads during the time of the experiment (Fig. 5A). In month six, spawned oocytes were found during a water change on one occasion. Moreover, specimens bearing gonads at winter conditions fragmented in a few cases, which did not happen at summer conditions. At winter conditions, there is no significant difference in length between specimens bearing gonads (sexually mature) and those without visible gonads (immature) (Mann–Whitney U test, W = 1451, p = 0.05), whereas mature specimens are significantly longer at summer conditions than immature ones (Mann–Whitney U test, W = 563.5, p < 0.05) (Fig. 5B, C).

Sexual maturation of Lineus sanguineus in the Bergen, Norway population. A Amount of specimens bearing gonads at summer (18 °C, 16 h light) and winter conditions (9 °C, 8 h light), examined at the beginning, halfway through, and at the end of the experiments. Number of specimens bearing gonads given in percent. Asterisk denotes presence of egg clutches. B At summer conditions there is a significant length difference of sexually mature and immature specimens. C At winter conditions, no significant length difference between sexually mature and immature specimens is present. Significance was tested based on the Wilcoxon rank sum test (***p < 0.001)
So far, no larva has been recorded for L. sanguineus. Here, we give the first incidental report on a pilidium larva in this species. Few larvae of L. sanguineus were found in the container with animals that had been sampled in Roscoff, France. The larvae appear to be typical heteronemertean pilidium larvae resembling the larvae of Riseriellus occultus (Beckers et al. 2015). The outer appearance is roughly similar to a 3 to 4-day old larva of R. occulus with an apical plate and an apical tuft, two lateral lappets with ciliated bands and a developed, blind-ending midgut (Fig. 6). The size of midgut and esophagus indicate beginning malnutrition as no appropriate food (such as unicellular planktonic algae) had been available.

Two young pilidium larvae (three to four days post fertilization) of Lineus sanguineus, differential interference contrast (A & B). The larval umbrella and the lateral lappets (la) are differentiating. An apical tuft (at) arises from the apical plate (ap). Esophagus (es) and sac-like midgut (mg) are developed, but the larvae did not take up any food. Note that the larvae resemble pilidium larvae of the closely related lineid species Riseriellus occultus (cf. Beckers et al. 2015)
Discussion
A rare case of cosmopolitan distribution in Lineus sanguineus
The results of our phylogenetic analysis clearly show that all specimens of L. sanguineus included in this study belong to only one species with a wide geographic range. Thus, as already previously shown, a division into four species based on geographic locality can again be refuted (Bierne et al. 1993; Riser 1993; Kang et al. 2015). Analogously to polychaete species, in which it has been stated that most species naturally have a relatively limited range of distribution (Hutchings and Kupriyanova 2018), a cosmopolitan distribution in a nemertean species currently represents an exception. On the contrary, cryptic speciation, i.e. a separation into several species, even within smaller areas, is a common phenomenon in Nemertea (e.g. Leasi et al. 2016 for Ototyphlonemertes, Krämer et al. 2017 for Lineus viridis and Lineus clandestinus, or Sagorny et al. 2019 for European Cephalothrix species). Nevertheless, a recent study in the fissiparous polychaete Proscoloplos cygnochaetus Day, 1954 showed that most likely three nominal species from different, distant localities actually represent one species that was assumed to have been transmitted from its native area of distribution in Europe to Australia and South Africa in the fouling community of ship hulls (Kelaher and Rouse 2003; Meyer et al. 2008). The situation in P. cygnochaetus superficially resembles that in L. sanguineus.
Like P. cygnochaetus, L. sanguineus is a species that is also known to show strong regenerative abilities enabling asexual reproduction by fissiparity. Since only little is known about sexual reproduction in this species, fissiparity was assumed to be the main mode of reproduction (Gontcharoff 1951; Bierne 1970; Gibson 1972). Therefore, it has been suggested that the cosmopolitan distribution of L. sanguineus is mainly the result of dispersal via anthropogenic means. (Riser 1993, 1994; Caplins and Turbeville 2011; Kang et al. 2015). Whereas nemertean species have already been found rafting on macroalgae (Thiel and Gutow 2005), the century-old importance and extent of cosmopolitan shipping traffic makes dispersal on ship hulls appear to be the more important vector for adult specimens (Kang et al. 2015). Due to their ability to establish new colonies from only one individual, population expansion after anthropogenic dispersal is likely more successful in asexually reproducing species (Waters and Roy 2003; Lázaro et al. 2009). For that reason, dispersal of adult specimens or fragments by ships, leading to multiple foreign introductions, has been regarded as so far best explanation for the cosmopolitan distribution of L. sanguineus (Riser 1994; Kang et al. 2015). Based on the frequency and extent of global shipping transport and the biology of Lineus sanguineus this mode of dispersal probably accounts for at least some of the global distribution of the species (Riser 1994; Runnels 2013). The formation of cysts, fragmentation and a habitat preference of fouling community might facilitate dispersal in the fouling community of ships (Riser 1994; Runnels 2013; Kang et al. 2015). Since several shipping routes, especially between northern Europe, the Atlantic coasts of North America, and the Indian Ocean have already been extensively traveled since the seventeenth century, gene flow based on anthropogenic dispersal could have been happening for centuries (Runnels 2013; Druzin 2016). Nevertheless, as already stated by Kang et al. (2015) some patterns of population distribution cannot be explained by frequent dispersal by ship.
If specimens of L. sanguineus were dispersed by anthropogenic means, one would expect more genetic variation in the native population whereas only a fraction of the native haplotypes should be found in the introduced population as shown for Cephalothrix simula, the currently most prominently documented nemertean example for anthropogenic introduction (Fernández-Álvarez and Machordom 2013; Kajihara et al. 2013; Faasse and Turbeville 2015; Sagorny et al. 2019). A pattern like this could not be detected in our datasets. Therefore, we argue that anthropogenic means most likely contribute to the cosmopolitan distribution, but other factors like rafting or dispersal of larvae cannot be ruled out by the results presented in this study. Furthermore, the genetic difference found in Mediterranean specimens of L. sanguineus is especially pronounced. This difference might lend evidence against anthropogenic dispersal on ship hulls, since the Mediterranean is traversed by one of the most-frequented freight routes worldwide. To elucidate the species distribution patterns further specimens from different localities need to be sampled and the biodiversity of the ship fouling communities should be investigated (Runnels 2013; Kang et al. 2015).
No restriction of gene-flow was found between populations of Lineus sanguineus
Whereas anthropogenic introduction and establishment of populations by asexual reproduction has been convincingly shown in P. cygnochaetus, the situation seems to be different in L. sanguineus. In truly asexual species one would expect only little genetic variation within populations because all specimens derive from the same founder individual and no recombination takes place (Johnson and Threlfall 1987; Bürger 1999). Additionally, high levels of genetic differentiation are expected between different populations as mutations happening in one population are maintained in this population but not shared with other populations (Palumbi 1992; Bürger 1999). However, this study shows that there is a high level of genetic variation within the species, but not between the populations. According to the haplotype networks generated based on the two different mitochondrial gene fragments (COI and 16S) haplotypes are shared by numerous individuals that originate from distant populations. This may be the consequence of using mitochondrial markers as these are often not sufficient to reliably differentiate between recently diverged populations. Therefore, we included a nuclear, more variable gene region in the form of the internal transcribed spacer (ITS). The haplotype network based on the ITS gene fragment shows more structuring, i.e. more haplotypes shared by less individuals. Nevertheless, there is no clear distinction between populations from different European collection sites. Most haplotypes that are shared by more than one specimen are found in specimens from more than one sampling site. Inferring our result from only three gene regions may limit its informative value. Nevertheless, the low levels of population divergence found especially in the ITS region are supported by the results of the genome-wide study by Ament-Velásquez et al. (2016). The observed haplotype distributions indicate that genetic exchange has to take place between distant populations, a conclusion that is also supported by the results of the AMOVA analysis of the mitochondrial gene fragments. Most variation is found between individuals within the same populations and there is only little structuring among regions, indicating that isolation by distance may not play a major part. This result is in contrast to the results of Runnels (2013), that imply isolation by distance, especially between different north-west Atlantic populations. Unfortunately, there are only few sequences available for some of the regional populations. To clarify the apparent contradiction between the published results and the data herein, more marker-sequences, especially from nuclear genes, should be included in future analyses. Herein, the only region that differs significantly from all other regions is the Mediterranean. The presence of the relatively narrow Strait of Gibraltar could account for a limited dispersal and genetic exchange between populations from the Mediterranean and Atlantic populations but more data is needed to resolve this question.
The observed population structuring contradicts the pattern expected for truly asexual species, therefore sexual reproduction has to occur to some extent. This structuring supports the hypothesis that fissiparity is the main mode of reproduction in L. sanguineus. Since only few sexually derived individuals per generation are enough to establish a genetic population structure comparable to fully sexual species (Bengtsson 2003; Hartfield et al. 2016), it could be possible that sexual reproduction is very rare in L. sanguineus and has therefore not been documented previously. As mentioned above, anthropogenic dispersal might also account for some of the observed haplotype structuring. The high differentiation between Norwegian and Canadian, and between Chilean and northern Atlantic populations could be the result of only infrequent dispersal by ship as these localities are not on main shipping routes (Rodrigue 2016). To fully answer this question, more data is needed on the correlation between population structuring and main shipping routes.
In contrast to the polychaete P. cygnochaetus (Meyer et al. 2008), neither recent anthropogenic introductions by hull fouling, nor subsequent establishment of populations exclusively by asexual reproduction could be substantiated in L. sanguineus by our data. Instead, our results indicate sexual reproduction resulting in some gene-flow between different populations. A population structure resulting from panmixia is also indicated by the analysis of genomic data of L. sanguineus, where no geographic structuring but normal levels of heterozygosity were observed (Ament-Velásquez et al. (2016). Therefore, we assume that at least the distribution and genetic variability of European populations are the result of at least infrequent sexual reproduction and thus larval dispersal leading to one large European metapopulation.
Although all data hint at successful sexual reproduction resulting in viable offspring, it has never been documented in L. sanguineus. In the French population studied by Gontcharoff (1951) only 1/3 of all specimens reached sexual maturity. In this study, we were able to obtain a few young pilidium larvae of L. sanguineus. As typical of heteronemertean larvae between three and four days, an apical tuft and lateral lappets were developed, as well as the midgut (Maslakova 2010; Beckers et al. 2015). The larvae closely resembled larvae of the heteronemertean species Riseriellus occultus Rogers, Junoy, Gibson and Thorpe 1993 (Beckers et al. 2015). Interestingly, these larvae occurred in a French population, in which oogenesis has been reported to be abortive (Gontcharoff 1951) The occurrence of larvae in L. sanguineus raised the question which factors influence gonad maturation and sexual reproduction. Although the larvae were not subjected to COI-barcoding, and thus direct evidence for their specific identity is absent, there is circumstantial evidence indicating a high likelihood of the larvae belonging to L. sanguineus. Only one other species that possesses superficially similar-looking larvae and has adults that could be taken for L. sanguineus is known to be present in the same habitat. However, this species, R. occultus has so far not been recorded from Roscoff. It is therefore unlikely that specimens of R. occultus had been misidentified as L. sanguineus. The COI-fragments taken from adult specimens collected at that occasion identify them as L. sanguineus. Furthermore, the reproductive period of R. occultus ranges from September to October, with gonopores being visible no earlier than late July (Beckers et al. 2015). Although gametogenesis starts as early as late March, the short time (5 weeks) the animals had spent at an elevated temperature (18 °C) in captivity seems to not be long enough, even for accelerated gonad maturity due to elevated temperatures. Therefore, even if specimens of R. occultus were erroneously present in the container from which the larvae were extracted, it would be very improbable that they had developed fertile gametes at the time when the larvae were found. Although we are confident about the identity of the larvae as being larvae of L. sanguineus, we admit that confirmation by DNA barcoding on a future occasion would be preferable, as it provides more straightforward, direct evidence.
Fissiparity as the prevalent reproductive mode in L. sanguineus
According to literature, the main factor influencing fissiparity in L. sanguineus is temperature (Coe 1930a). Former studies have shown that high temperatures promote fragmentation and subsequent regeneration. Therefore, asexual reproduction mainly takes place during the summer months (Coe 1929, 1930a; Gontcharoff 1951; Reutter 1967). In general, lower temperatures decelerate the rate of fragmentation down to almost complete inhibition at temperatures between 5 and 10 °C, whereas temperatures around 21 °C yield the highest number of fragments and fastest regeneration (Coe 1930a; Reutter 1967). For this reason, we chose temperatures that range at the respective ends of this spectrum (low: 9 °C; high: 18 °C) for our experiments. In contrast to published data, the specimens from the Bergen population fragmented to a higher degree at winter temperatures. Unexpectedly, even some sexually mature specimens fragmented under winter conditions although fragmentation was reported to be strongly reduced and regeneration to be less complete in sexually mature specimens (Coe 1930a; Gontcharoff 1951; Reutter 1967).
With regard to the influence of photoperiod on fragmentation, our results do not contradict the findings of Coe (1930a) and Reutter (1967) that temperature seems to have a stronger effect on fragmentation than light. However, photoperiod seems to have an influence on regeneration and growth of the animals. Our results might indirectly support the findings of Arnould and Vernet (1995) that regeneration is negatively correlated with melatonin production. Long hours without light usually lead to the production of melatonin in the closely related species Lineus lacteus (Arnoult et al. 1994). Melatonin retards or even completely inhibits regeneration in Lineus sanguineus, depending on the concentration, possibly by reducing mitotic activity (Arnoult and Vernet 1995). These assumptions suggest that longer hours of light should in turn lead to more regenerating and also longer individuals. This expectation is met for the Bergen specimens where summer conditions resulted in more regenerating fragments as well as faster growing specimens. However, elucidating the exact physiological mechanisms underlying fragmentation and regeneration in L. sanguineus exposed to changing light and/or temperature needs further investigation.
Sexual reproduction—more frequent than previously assumed
The effects of light and temperature on sexual maturation on L. sanguineus are not well-studied. Winter conditions have for long been assumed to trigger egg maturation and sexual reproduction in the fissiparous species Lineus sanguineus (Coe 1899; Gibson 1972; Gontcharoff 1951; McIntosh 1873–1874; Riser 1994). According to Gontcharoff (1951) oogenesis is abortive in L. sanguineus in the majority of female specimens from Brittany, whereas in males, normal spermatogenesis occurs (Riser 1994). In October, testes and ovaries are formed between the midgut diverticula (McIntosh 1873–1874; Gontcharoff 1951; Bierne 1983). From January to March the testes are filled with small, awl-handle shaped, mobile spermatozoa that resemble spermatozoa of Lineus lacteus and Riseriellus occultus (McIntosh 1873–1874; Gontcharoff 1951; Döhren et al. 2010; Beckers et al. 2015). At the same time of the year, numerous small, mature oocytes can be observed in female specimens (McIntosh 1873–1874; Riser 1994). Oocytes have been described to undergo autolysis or are resorbed into the intestine (Gontcharoff 1951; Riser 1994). Moreover, females of this species produce only very few oocytes per individual leading to rather small clutches of eggs that have been found rarely. Those eggs are shed directly into the water, lacking a gelatinous sheath, and have been observed to develop into swimming gastrulae (Coe 1899; Gontcharoff 1951; Bierne et al. 1993). Aside from this, nothing is known about the development of Lineus sanguineus although Coe (1899, 1943) states that L. sanguineus is well-suited for embryonic studies.
In the closely related species Lineus lacteus and Lineus ruber, more gonads develop under colder temperature than under warmer temperature regimes with the more southwardly distributed species Lineus lacteus developing more gonads at 12 °C than the more northwardly distributed species Lineus ruber (Vernet and Bierne 1988). In these species, a gonad-inhibiting hormone of unknown nature (GIH) produced in the cerebral ganglia was made responsible for gonad maturation (Vernet and Bierne 1988, 1993; Arnoult and Vernet 1996). During the summer months, synthesis of this factor is supposedly increased, leading to a sexual rest (Vernet and Bierne 1993; Arnoult and Vernet 1996). The amount of light apparently has no influence on gonadogenesis in L. lacteus and L. ruber (Vernet and Bierne 1988, 1993).
For the reasons stated above, we expected low temperatures to promote sexual maturity in L. sanguineus. In the specimens from Norway, some influence of temperature could be shown as winter conditions led to a slower decrease in the number of mature specimens (measured by the presence of gonopores) and even the deposition of eggs. Thus, maintenance of a sexually mature state was seen in more specimens at the lower temperature (9 °C) than at the higher temperature (18 °C). This potentially indicates a connection between temperature and sexual reproduction in L. sanguineus. The reason why the number of specimens to show gonads didn’t rise at lower temperatures can only be speculated about. Since the majority of specimens from the Norway population already had gonads at the onset of the experimental procedure, it is conceivable that spawning in several specimens might not have been noticed and subsequently the gonads were resorbed. Unfortunately, the potential influence of light on gonad maturation could not be directly assessed since the number of Norwegian specimens had been too small from the beginning to set up conditions that uncouple temperature and light exposition (i.e. long photoperiod with low temperature versus short photoperiod and higher temperature) with a reasonable sample size. Successful gonad maturation was also observed in specimens from Uruguay (Sivaradjam and Bierne 1981; Bierne 1983; Tarpin and Bierne 1995). Therefore, it might be possible that populations from colder habitats like Norway and Uruguay are more likely to develop gonads and reproduce sexually than populations from warmer habitats. To confirm this hypothesis, specimens from localities with warmer water temperatures will need to be experimentally tested.
Conclusions and outlook
Our results show that Lineus sanguineus is a cosmopolitan species. Although it has been reported that it mainly relies on asexual reproduction by fissiparity, L. sanguineus is able to reproduce sexually to promote genetic exchange. One question that prevails is whether the main reason for the cosmopolitan distribution is a result of larval dispersal, adult rafting or anthropogenic dispersal, e.g. along international freight routes. Of focal interest in this regard is the genetically deviating population found in the Mediterranean.
In the specimens from a population from Norway, sexual maturation seems to be result of low temperatures and/or short light period, but is only reached in approximately half of the specimens. Seasonal changes in day-length and water temperature therefore are apparently not the only factors influencing sexual reproduction in L. sanguineus. It has to be investigated whether these different reactions are lineage specific or influenced by population density, food provisioning, local adaptation or epigenetic factors. Another aspect of future research regards the interplay between sexual and asexual reproduction and its ecological consequences for L. sanguineus as an upcoming spiralian model species to study the different (sexual vs. asexual) reproductive modes.
Data availability
All sequence data have been deposited in the GenBank database (accession numbers provided in Table 1). Detailed results of the experimental setups are provided in the electronic supplementary files.
References
Ament-Velásquez SL, Figuet E, Ballenghien M, Zattara EE, Norenburg JL, Fernández-Álvarez FA, Bierne J, Bierne N, Galtier N (2016) Population genomics of sexual and asexual lineages in fissiparous ribbon worms (Lineus, Nemertea): hybridization, polyploidy and the Meselson effect. Mol Ecol 25(14):3356–3369. https://doi.org/10.1111/mec.13717
Andrade SCS, Strand M, Schwartz M, Chen H, Kajihara H, von Döhren J, Sun S, Junoy J, Thiel M, Norenburg JL, Turbeville JM, Giribet G, Sundberg P (2012) Disentangling ribbon worm relationships: multi-locus analysis supports traditional classification of the phylum Nemertea. Cladistics 28(2):141–159. https://doi.org/10.1111/j.1096-0031.2011.00376.x
Arnoult F, Vernet G (1995) Inhibition of regeneration by melatonin in nemertean worms of the genus Lineus. Comp Biochem Physiol 110(4):319–328. https://doi.org/10.1016/0300-9629(94)00181-R
Arnoult F, Vernet G (1996) Influence of melatonin on gonad maturation in Lineus lacteus (Heteronemertini, Lineidae). J Exp Zool 274(2):138–143. https://doi.org/10.1002/(SICI)1097-010X(19960201)274:2%3c138:AID-JEZ7%3e3.0.CO;2-Q
Arnoult F, Vivien-Roels B, Pévet P, Vernet G (1994) Melatonin in the Nemertine Worm Lineus lacteus: Identification and Daily Variations. Neurosignals 3(6):296–301. https://doi.org/10.1159/000109557
Avise JC (2000) Phylogeography: the history and formation of species. Harvard University Press, Cambridge
Beckers P, Bartolomaeus T, von Döhren J (2015) Observations and experiments on the biology and life history of Riseriellus occultus (Heteronemertea: Lineidae). Zool Sci 32(6):531–546. https://doi.org/10.2108/zs140270
Bely AE, Nyberg KG (2010) Evolution of animal regeneration: re-emergence of a field. Trends Ecol Evol 25(3):161–170. https://doi.org/10.1016/j.tree.2009.08.005
Bengtsson BO (2003) Genetic variation in organisms with sexual and asexual reproduction. J Evol Biol 16(2):189–199. https://doi.org/10.1046/j.1420-9101.2003.00523.x
Bierne J (1990) Lineus as a model for studying developmental processes. Int J Dev Biol 34:245–253
Bierne J, Tarpin M, Vernet G (1993) A reassessment of the systematics and a proposal for the phylogeny of some cosmopolitan Lineus species (Nemertea). Hydrobiologia 266:159–168. https://doi.org/10.1007/978-94-011-2052-4_11
Bierne J (1970) Recherches sur la différenciation sexuelle au cours de l'ontogenèse et de la régénération chez le némertien Lineus ruber (Müller). Doctoral dissertation
Bierne J (1983) Nemertina. In: Adiyodi KG, Adiyodi RG (Eds) Reproductive biology of invertebrates.: Volume I: oogenesis, oviposition, and oosorption, vol 1. John Wiley, Chichester, pp 147–167
Bürger R (1999) Evolution of genetic variability and the advantage of sex and recombination in changing environments. Genetics 153(2):1055
Cahill AE, de Jode A, Dubois S, Bouzaza Z, Aurelle D, Boissin E, Chabrol O, David R, Egea E, Ledoux J-B, Mérigot B, Weber AA-T, Chenuil A (2017) A multispecies approach reveals hot spots and cold spots of diversity and connectivity in invertebrate species with contrasting dispersal modes. Mol Ecol. https://doi.org/10.1111/mec.14389
Caplins SA, Turbeville JM (2011) The occurrence of Ramphogordius sanguineus (Nemertea, Heteronemertea) in the intertidal zone of the atlantic coast of virginia and new observations on its feeding behavior. Banisteria 38:65–70
Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol 17(4):540–552. https://doi.org/10.1093/oxfordjournals.molbev.a026334
Clement M, Posada D, Crandall KA (2000) TCS: a computer program to estimate gene genealogies. Mol Ecol 9(10):1657–1659. https://doi.org/10.1046/j.1365-294x.2000.01020.x
Coe WR (1899) Notes on the times of breeding of some common New England nemerteans. Science 9(214):167–169. https://doi.org/10.1126/science.9.214.167
Coe WR (1929) Regeneration in nemerteans. J Exp Zool 54:411–459
Coe WR (1930a) Asexual reproduction in nemerteans. Physiol Zool 3(3):297–308
Coe WR (1930b) Regeneration in nemerteans. J Exp Zool. https://doi.org/10.1017/CBO9781139923651.027
Coe WR (1934) Analysis of the regenerative processes in nemerteans. Biol Bull 66:304–3015
Coe WR (1943) Biology of the nemerteans of the atlantic coast of North America. Trans Conn Acad Arts Sci 35:129–328
Dawson MN, Jacobs DK (2001) Molecular evidence for cryptic species of Aurelia aurita (Cnidaria, Scyphozoa). Biol Bull 200(1):92–96. https://doi.org/10.2307/1543089
Druzin B (2016) Towards a theory of spontaneous legal standardization. J Int Disp Settl 8(3):403–431. https://doi.org/10.1093/jnlids/idw023
Excoffier L, Smouse PE, Quattro JM (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131(2):479
Faasse MA, Turbeville JM (2015) The first record of the north-west Pacific nemertean Cephalothrix simula in northern Europe. Mar Biodivers Rec 8:515. https://doi.org/10.1017/S1755267214001523
Fernández-Álvarez FÁ, Machordom A (2013) DNA barcoding reveals a cryptic nemertean invasion in Atlantic and Mediterranean waters. Helgol Mar Res 67(3):599–605. https://doi.org/10.1007/s10152-013-0346-3
Folmer O, Black M, Hoeh W, Lutz R, Vrijenhoek R (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol Mar Biol Biotech 3:294–299
Gibson R (1972) Nemerteans. Hutchinson & Co, London
Gibson R (1982) British Nemerteans, vol 24. Cambridge University Press, Cambridge
Gibson R (1995) Nemertean genera and species of the world: an annotated checklist of original names and description citations, synonyms, current taxonomic status, habitats and recorded zoogeographic distribution. J Nat Hist 29(2):271–561. https://doi.org/10.1080/00222939500770161
Gómez A, Serra M, Carvalho GR, Lunt DH (2002) Speciation in ancient cryptic species complexes: evidence from the molecular phylogeny of Brachionus plicatilis (Rotifera). Evolution 56(7):1431–1444. https://doi.org/10.1111/j.0014-3820.2002.tb01455.x
Gontcharoff M (1951) Biologie de la régénération et de la reproduction chez quelques Lineidae de France. Ann Des Sc Nat Zool: 151–235
Hall TA (Ed) (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT, 41. Oxford University Press
Hartfield M, Wright SI, Agrawal AF (2016) Coalescent times and patterns of genetic diversity in species with facultative sex: effects of gene conversion, population structure, and heterogeneity. Genetics 202(1):297–312. https://doi.org/10.1534/genetics.115.178004
Hasegawa M, Kishino H, Yano T (1985) Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J Mol Evol 22(2):160–174. https://doi.org/10.1007/BF02101694
Hiebert TC, Hunt M (2015) Ramphogordius sanguineus. In: Hiebert TC, Butler BA, Shanks AL (eds) Oregon estuarine invertebrates: Rudys' illustrated guide to common species, 3rd Edn. University of Oregon Libraries, Oregon
Hutchings P, Kupriyanova E (2018) Cosmopolitan polychaetes – fact or fiction? Personal and historical perspectives. Invertebr Syst 32(1):1–9
Ikenaga J, Hookabe N, Kohtsuka H, Yoshida M, Kajihara H (2019) A Population Without Females: Males of Baseodiscus delineatus (Nemertea: Heteronemertea) Reproduce Asexually by Fragmentation. Zool Sci 36(4):348–353
Jablonski D (1986) Larval ecology and macroevolution in marine invertebrates. Bull Mar Sci 39(2):565–587
Johnson MS, Threlfall TJ (1987) Fissiparity and population genetics of Coscinasterias calamaria. Mar Biol 93:517–525. https://doi.org/10.1007/BF00392789
Kajihara H, Sun S-C, Chernyshev AV, Chen H, Ito K, Asakawa M, Maslakova SA, Norenburg JL, Strand M, Sundberg P, Iwata F (2013) Taxonomic identity of a tetrodotoxin-accumulating ribbon-worm cephalothrix simula (nemertea: palaeonemertea): a species artificially introduced from the Pacific to Europe. Zool Sci 30(11):985–997
Kang X-X, Fernández-Álvarez FÁ, Alfaya JEF, Machordom A, Strand M, Sundberg P, Sun S-C (2015) Species diversity of Ramphogordius sanguineus/Lineus ruber-Like Nemerteans (Nemertea: Heteronemertea) and Geographic Distribution of R. sanguineus. Zool Sci 32(6):579–589. https://doi.org/10.2108/zs150064
Katoh K, Rozewicki J, Yamada KD (2019) MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform 20(4):1160–1166. https://doi.org/10.1093/bib/bbx108
Kelaher BP, Rouse GW (2003) The role of colonization in determining spatial patterns of Proscoloplos bondi sp. Nov. (Orbiniidae: Annelida) in coralline algal turf. Mar Biol 143(5):909–917. https://doi.org/10.1007/s00227-003-1143-3
Krämer D, Schmidt C, Podsiadlowski L, Beckers P, Horn L, von Döhren J (2017) Unravelling the Lineus ruber/viridis species complex (Nemertea, Heteronemertea). Zool Scr 46(1):111–126. https://doi.org/10.1111/zsc.12185
Lázaro EM, Sluys R, Pala M, Stocchino GA, Baguñà J, Riutort M (2009) Molecular barcoding and phylogeography of sexual and asexual freshwater planarians of the genus Dugesia in the Western Mediterranean (Platyhelminthes, Tricladida, Dugesiidae). Mol Phylogenet Evol 52(3):835–845. https://doi.org/10.1016/j.ympev.2009.04.022
Leasi F, Andrade SCdS, Norenburg J (2016) At least some meiofaunal species are not everywhere. Indication of geographic, ecological and geological barriers affecting the dispersion of species of Ototyphlonemertes (Nemertea, Hoplonemertea). Mol Ecol 25(6):1381–1397. https://doi.org/10.1111/mec.13568
Mantel N (1967) The detection of disease clustering and a generalized regression approach. Cancer Res 27(2 Part 1):209
Maslakova SA (2010) Development to metamorphosis of the nemertean pilidium larva. Front Zool 7(1):30. https://doi.org/10.1186/1742-9994-7-30
McIntosh WC (1873–1874) A monograph of the British marine annelids: The Nemerteans. Ray Society, London
Meyer A, Bleidorn C, Rouse GW, Hausen H (2008) Morphological and molecular data suggest a cosmopolitan distribution of the polychaete Proscoloplos cygnochaetus Day, 1954 (Annelida, Orbiniidae). Mar Biol 153(5):879–889. https://doi.org/10.1007/s00227-007-0860-4
Nei M, Kumar S (2000) Molecular evolution and phylogenetics. Oxford University Press
Nylander JAA (2004) MrModeltest v2. Program distributed by the author
Palumbi SR (1992) Marine speciation on a small planet. Trends Ecol Evol 7(4):114–118. https://doi.org/10.1016/0169-5347(92)90144-Z
Palumbi SR, Grabowsky G, Duda T, Geyer L, Tachino N (1997) Speciation and population genetic structure in tropical Pacific sea urchins. Evolution 51(5):1506–1517. https://doi.org/10.1111/j.1558-5646.1997.tb01474.x
Palumbi SR, Martin A, Romano S, McMillan WO, Stice L, Grabowski G (1991) The simple fool's guide to PCR, ver. 2.0. Dept. of Zoology and Kewalo Marine Laboratory, Hawaii
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in excel. Population genetic software for teaching and research. Mol Ecol Notes 6(1):288–295. https://doi.org/10.1111/j.1471-8286.2005.01155.x
Peakall R, Smouse PE (2012) GenAlEx 6.5: genetic analysis in excel. Population genetic software for teaching and research–an update. Bioinformatics 28(19):2537–2539. https://doi.org/10.1093/bioinformatics/bts460
R Core Team (2019) R: A language and environment for statistical computing
Reutter K (1967) Untersuchungen zur ungeschlechtlichen Fortpflanzung und zum Regenerationsvermögen von Lineus sanguineus Rathke (Nemertini). Wilhelm Roux Arch Entwickl Mech Org 159(2):141–202. https://doi.org/10.1007/BF00573439
Riser NW (1993) Observations on the morphology of some North American nemertines with consequent taxonomic changes and a reassessment of the architectonics of the phylum. In: Gibson R, Moore J, Sundberg P (eds) Advances in Nemertean Biology. Springer, Netherlands, Dordrecht, pp 141–157
Riser NW (1994) The morphology and generic relationships of some fissiparous heteronemertines. Proc Biol Soc Wash 107(3):548–556
Rodrigue J-P (2016) Maritime transport. In: Richardson D, Castree N, Goodchild MF, Kobayashi A, Liu W, Marston RA (eds) International encyclopedia of geography: people, the Earth, environment and technology. Wiley, Oxford, pp 1–7
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, Sánchez-Gracia A (2017) DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol 34(12):3299–3302. https://doi.org/10.1093/molbev/msx248
Runnels C (2013) Phylogeography and species status of Ramphogordius sanguineus. Master Thesis, Virginia Commonwealth University
Sagorny C, Wesseler C, Krämer D, von Döhren J (2019) Assessing the diversity and distribution of Cephalothrix species (Nemertea: Palaeonemertea) in European waters by comparing different species delimitation methods. J Zool Syst Evol Res 57(3):497–519. https://doi.org/10.1111/jzs.12266
Sanger F, Nicklen S, Coulson AR (1977) DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci USA 74(12):5463–5467. https://doi.org/10.1073/pnas.74.12.5463
Sivaradjam S, Bierne J (1981) Sex differentiation in bilaterally allophenic animals produced by cloning of two bipartite male/female chimaeras of Lineus sanguineus. J Embryol Exp Morphol 65:173–184
Sundberg P, Saur M (1998) Molecular phylogeny of some European heteronemertean (Nemertea) species and the monophyletic status of Riseriellus, Lineus, and Micrura. Mol Phylogenet Evol 10(3):271–280. https://doi.org/10.1006/mpev.1998.0543
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30(12):2725–2729. https://doi.org/10.1093/molbev/mst197
Tarpin M, Bierne J (1995) Expression of sex-specific molecular markers in clones of bipartite allophenic nemertines produced by somatic embryogenesis from Lineus sanguineus male/female chimera fragments. Int J Dev Biol 39:415–420
Templeton AR, Crandall KA, Sing CF (1992) A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 132(2):619–633
Thiel M, Gutow L (2005) The ecology of rafting in the marine environment. II. The rafting organisms and community. In: Oceanography and Marine Biology. CRC Press, pp 289–428
Thollesson M, Norenburg JL (2003) Ribbon worm relationships: a phylogeny of the phylum Nemertea. Proc Royal Soc B 270(1513):407–415. https://doi.org/10.1098/rspb.2002.2254
Vernet G, Bierne J (1988) Neuroendocrine control of gonadogenesis in regenerating Lineus lacteus (Heteronemertea). Hydrobiologia 156(1):53–60. https://doi.org/10.1007/BF00027978
Vernet G, Bierne J (1993) The influence of light and sea water temperature on the reproductive cycle of Lineus ruber (Heteronemertea). Hydrobiologia 266(1):267–271. https://doi.org/10.1007/BF00013374
von Döhren J, Beckers P, Vogeler R, Bartolomaeus T (2010) Comparative sperm ultrastructure in Nemertea. J Morphol 271(7):793–813. https://doi.org/10.1002/jmor.10834
Waters J, Roy M (2003) Global phylogeography of the fissiparous sea-star genus Coscinasterias. Mar Biol 142(1):185–191. https://doi.org/10.1007/s00227-002-0915-5
Wickham H (2016) ggplot2: elegant graphics for data analysis. Springer
Zattara EE, Fernandez-Alvarez FA, Hiebert TC, Bely AE, Norenburg JL (2019) A phylum-wide survey reveals multiple independent gains of head regeneration ability in Nemertea. Proc Royal Soc B. https://doi.org/10.1098/rspb.2018.2524
Acknowledgements
The authors thank Christiane Todt and Henrik Glenner for the invitation to the Marine Biological Station Espegrend in Bergen and the staff of the station for facilitating sample collection. We also would like to gratefully acknowledge Prof. Dr. Andreas Hejnol for collecting samples from Bergen and the staffs of the Station Marine de Wimereux, the Station Biologique de Roscoff and the Station de Biologie Marine et Marinarium de Concarneau in France, as well as the Wattenmeerstation in List auf Sylt and the Biologische Anstalt Helgoland of the Alfred-Wegener-Institute (AWI) for providing facilities and accommodation during several collecting trips.
Funding
Open Access funding enabled and organized by Projekt DEAL. Not applicable.
Author information
Authors and Affiliations
Contributions
Both authors contributed to the study conception and design. Collection of genetic data, formal analysis and visualization were performed by CS. Experimental procedures were executed by JvD. The first draft of the manuscript was written by CS with additional remarks by JvD. All authors read, revised and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethics approval
All applicable international, national, and institutional guidelines for sampling of organisms for this study have been followed.
Additional information
Responsible Editor: O. Puebla.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sagorny, C., von Döhren, J. Occasional sexual reproduction significantly affects the population structure of the widespread, predominantly asexually reproducing marine worm Lineus sanguineus (Nemertea: Pilidiophora). Mar Biol 169, 92 (2022). https://doi.org/10.1007/s00227-022-04077-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00227-022-04077-0