
Abstract
This investigation examines the source of the disparity between experimental values of the light scattering second virial coefficient \({A}_{2}\) (mL.mol/g2) for proteins and those predicted on the statistical mechanical basis of excluded volume. A much better theoretical description of published results for lysozyme is obtained by considering the experimental parameters to monitor the difference between the thermodynamic excluded volume term and its hydrodynamic counterpart. This involves a combination of parameters quantifying concentration dependence of the translational diffusion coefficient obtained from dynamic light scattering measurements. That finding is shown to account for observations of a strong correlation between \({A}_{2}{M}_{2}\) (mL/g), where M2 is the molar mass (molecular weight) of the macromolecule and the diffusion concentration parameter \({k}_{D}\) (mL/g). On the grounds that \({k}_{D}\) is regarded as a hydrodynamic parameter, the same status should be accorded the light scattering second virial coefficient rather than its current incorrect thermodynamic designation as \({B}_{2}\) (mL.mol/g2), or just B, the osmotic second virial coefficient for protein self-interaction.
Similar content being viewed by others
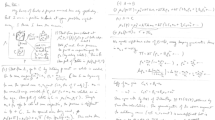


Introduction
In accordance with standard textbook doctrine (Tanford 1961; Cantor and Schimmel 1980; Van Holde 1985) the nonideality parameter \({A}_{2}\) (mL.mol.g−2) determined from the linear concentration dependence of “classical” or “total intensity” light scattering intensity measurements for protein solutions has routinely been identified with \({B}_{2},\) the osmotic second virial coefficient (mL mol \({\mathrm{g}}^{-2}\)) for solute self-interaction. However, that designation was called into question (Deszczynski et al. 2006; Winzor et al. 2007) as the result of reports of negative values (George and Wilson 1994; Muschol and Rosenberger 1995; Rosenbaum and Zukoski 1996)—a finding incompatible with the interpretation of thermodynamic nonideality on the statistical-mechanical basis of excluded volume (McMillan and Mayer 1945; Hill 1968). Experimental support for that contention has come from rigorous estimation of the osmotic second virial coefficient for protein self-interaction by sedimentation equilibrium (Wills and Winzor 1992; Wills et al. 1993), which has yielded positive values of \({B}_{2}\) for lysozyme (Behlke and Ristau 1999), equine serum albumin (Desczynski et al. 2006) and ovalbumin (Winzor et al. 2007) under comparable conditions to those yielding negative \({A}_{2}\) values from “static” light scattering measurements (Muschol and Rosenberger 1995; Guo et al. 1999; Winzor et al. 2007). The thermodynamic second virial coefficient B2 when measured correctly from osmotic pressure or sedimentation equilibrium in the analytical ultracentrifuge (Hall et al 1999) is useful in the description of the thermodynamic nonideality behaviour of macromolecular solutions and in the elucidation of macromolecular shape, conformation in solution and solvation (Rallison and Harding 1985; Harding 1985; Winzor et al 2001; Serdyuk et al 2007 and references cited therein).
On the grounds that the quantitative expressions derived from a more rigorous thermodynamic treatment of the concentration dependence of static light scattering experiments still failed to generate the negative \({A}_{2 }\) versus c2 dependence (Winzor et al. 2007), we now explore the possibility that the linear concentration coefficient obtained from light scattering intensity measurements should be reclassified as a hydrodynamic steady-state parameter (Winzor et al. 2007).
Theoretical considerations
Caution is required when defining the second virial coefficient for a macromolecular solution because of the different conditions under which the solute concentration can be varied (Winzor and Wills 1994). Theory has traditionally been written for the condition of osmotic equilibrium, which is a convenient standard to which changes in thermodynamic quantities can be referred. Choice of this standard condition then allows the establishment of a formal equivalence between the theory of imperfect gases (van der Waals 1873) and nonideal macromolecular solutions (Hill 1959; Wills and Winzor 2005).
Macromolecular solutions: thermodynamic considerations
The inclusion of a single protein solute (species 2) in solvent (species 1) at constant temperature gives rise to one of two situations. In classical osmometry, for example, the chemical potential of solvent (\(\mu_{1}\) ) in the protein-containing phase (α) and solvent phase (β) remains equal to that of solvent at atmospheric pressure; and hence complies with the condition of osmotic equilibrium discussed above. Because buffer components and additional small cosolutes are also in partition equilibrium between the two phases, they too can be regarded as part of the solvent, whereupon the solution becomes a single-solute system. A totally different situation arises in situations (as in light scattering measurements) where constant pressure P is the second constraint. Because the osmotic equilibrium condition is not met: the solvent chemical potential then becomes dependent upon the protein concentration. Consequently, separate definitions of the solute chemical potential (\(\mu_{2}\)) are required for the two situations (Hill 1959).
Under the constraints of constant temperature and solvent chemical potential the thermodynamic activity of the protein \({(z}_{2})\) is related to its molar concentration (\({C}_{2}\)) by the relationship (Hill 1959; Wills et al. 2015)
where the solute thermodynamic activity z2 is a molar quantity in the sense that it equals the molar concentration \({C}_{2}\) in the ideal limit of infinite dilution (a situation denoted by superscript o); and is therefore most appropriately expressed as the product of \({C}_{2}\) and a dimensionless activity coefficient \(\gamma_{2}\) . The osmotic pressure can be written in virial form as
where M2 is the molar mass of the solute (g.mol−1) and B2 is the second virial coefficient (mL.mol.g−2). Rewriting with the second virial coefficient B2 expressed in the dimensions of exclusion volume (mL.mol.−1) \(B_{22} = B_{2 }.M_{2}^{2}\)
These and other terms are summarised in Table 1.
Using a purely thermodynamic argument, Hill (1959) has shown that the expression
defines the molar activity coefficient. For a spherical solute with radius \(R_{2}\) his second virial coefficient can be expressed in terms of the potential-of-mean-force, \(u_{22}\), between two molecules separated by centre-to-centre distance r via the equations (McMillan and Mayer 1945; Mayer 1950; Hill 1968)
where \({\mathrm{k}}_{\mathrm{B}}\) is the Boltzmann constant; and where Avogadro’s number \(\left( {N_{A} } \right)\) converts the virial coefficient from a molecular to a molar basis. The first integral in Eq. (4a) accounts for the excluded volume for two uncharged spheres (the hard-sphere contribution \(B_{22}^{HS}\)), whereas the second integral accommodates the additional contribution of the perturbation of the chemical potential arising from electrostatic interaction \(\left( {B_{22}^{EL} } \right)\) via the function \(u_{22} \left( r \right)\) written in the form
for a protein bearing net charge \({Z}_{2}\) (not to be confused with the activity z2). The factor of 1000 reflects calculation of the Debye–Hückel inverse screening length κ (in cm−1) as 3.27 × \({10}^{7}\sqrt{{I}_{M}}\), where \({I}_{M}\) is the ionic strength recorded on the conventional molar scale (mol.L−1). Solution of Eq. (4) by approximating the Mayer function as
leads to the expression (Wills and Winzor 2009)
which establishes that the osmotic second virial coefficient for macromolecule self-interaction can only assume positive values because of convergence of the series of charge-dependent terms with alternating sign.
For solutions with temperature and pressure as fixed constraints, the thermodynamic activity of the macromolecule (\({a}_{2}\)) is defined (Hill 1968) by the equation
in which \({a}_{2}\) is a molal quantity and therefore most appropriately expressed as the product of molal concentration \({m}_{2}\) and a dimensionless activity coefficient \({y}_{2}\). Under these conditions the counterparts of Eqs. (2) and (3) for the change in solvent chemical potential due to the addition of solvent become (Hill 1968; Wills et al 1993)
where \({C}_{22}\) is the molal second virial coefficient for macromolecule self-interaction. Unlike its molar counterpart \({B}_{22}\), the molal second virial coefficient (\({C}_{22}\)) is not generally amenable to statistical-mechanical interpretation. However, the assumption of solution incompressibility (an acceptable approximation for aqueous systems) allows the expression of a molal concentration in terms of its molar counterpart as
where \({M}_{2}\) and \({\overline{v} }_{2}\) denote the molar mass and partial specific volume, respectively, of the protein, and where \(\gamma_{1}\) is the solvent density. It then follows that the two second virial coefficients are related by the expression (Wills et al. 1993)
which allows conversion of the molal second virial coefficient \({C}_{22}\) to its molar osmotic second virial counterpart \({B}_{22}\).
The above consideration of nonideality under the constraints of fixed temperature and pressure does, of course, refer to a single-solute system—a protein dissolved in solvent (water). As noted in the early days of static light scattering measurements (Kirkwood and Goldberg 1950; Stockmayer 1950) the monitoring of solute thermodynamic nonideality on the molal scale mandates the inclusion of additional virial coefficient terms for protein interaction with buffer components and other small cosolutes, which cannot be regarded as part of the solvent. It therefore becomes necessary to distinguish between nonideality reflecting the second virial coefficient for protein self-interaction (\({C}_{22}\)) and that emanating from physical protein–cosolute interaction (designated for simplicity of presentation as \({C}_{23}\)). The previous expression for the activity coefficient [Eq. (3)] needs modification to the form
to account for the physical interaction between protein and the cosolute present at molal concentration \({m}_{3}\).
More than fifty years elapsed before these requirements resurfaced in an investigation (Winzor et al. 2007) designed to test whether nondeality arising from protein–cosolute interactions could account for the negative light scattering second virial coefficients obtained in the presence of high concentrations of uncharged cosolutes, such as polyethylene glycol (Vivarés and Bonneté 2002) and sucrose (Winzor et al. (2007), for which nonideality reflects only the hard-sphere excluded volume interactions. Interest in this possibility was triggered by the presence of the term–\(\left( {C_{23} /\rho_{1} } \right)^{2}\) in the expression for the light scattering second virial coefficient (Kirkwood and Goldberg 1950; Winzor et al. 2007). Although that endeavour did lead to a decrease in the magnitude of \({A}_{2}\), the effect was minor compared to the experimentally observed dependence of \({A}_{2}\) upon \({C}_{3.}\) Furthermore, those calculations neglected the contribution of a third virial coefficient term, \(C_{223} /\rho_{2}^{2}\), which, it transpires, effectively counters that from\(\left( {C_{23} /\rho_{1} } \right)^{2}\). C223 is a parameter with the dimensions of a third virial coefficient reflecting the potential-of-mean-force interaction of a single cosolute molecule, which may be an electrolyte with a pair of protein molecules, and about which little is known, so it is effectively a fitting parameter (Deszczynski et al. 2006).
For present purposes we therefore proceed on the basis that the effects of thermodynamic nonideality in light scattering measurements on buffered aqueous protein solutions can be described adequately by single-solute theory (the standard practice). The respective expressions for m2 and C22 continue to be given by Eqs. 11 and 12, but with ρ1 the density of the cosolute-supplemented solvent.
Because the thermodynamic excluded volume parameter can only assume positive values, we also examine the possibility that the negative values of \({A}_{2}\) reported in the literature may include hydrodynamic contributions.
Macromolecular solutions: a hydrodynamic perspective
The first detailed consideration of the effect of hydrodynamic interactions on Brownian motion involving the net flux of solute was provided by Batchelor (1976) in the context of concentration-dependent diffusion in sedimentation velocity for solutions dilute enough that only pairwise interactions between particles were significant. In that study a combination of statistical-mechanical and hydrodynamic approaches led to description of the concentration dependence of the diffusion coefficient D for a rigid, uncharged spherical particle under such very dilute conditions as
in which \(\phi\), the volume fraction occupied by the diffusing particle = c2.vs, the product of the weight-concentration \({c}_{2}\) (g/ml) of the solute and its solvated specific volume (ml/g)\({\nu}_{s} = 4\pi N_{A} R_{2}^{3} /\left( {3M_{2} } \right)\) . \(\lambda_{T}\) and \(\lambda_{H}\) are coefficients representing the thermodynamic and hydrodynamic nonideality coefficients (the latter equivalent to Ks in Harding and Johnson (1985a, b)). \(D_{o} = \left( {{\text{k}}_{{\text{B}}} T} \right)/(6\pi \eta_{1} R_{2})\) is the “ideal” translational diffusion coefficient obtained experimentally in the limit of zero solute concentration. From Eq. (7) it is evident that the factor \({\lambda }_{T}=\) 8 in Eq. (14) corresponds to \(2{B}_{22}^{HS}/{M}_{2}\), the volume from which the centres of two uncharged solute molecules are mutually excluded; and is therefore a thermodynamic factor. Hydrodynamic factors are incorporated into the second term, \({\lambda }_{H}=\) 6.55 of Eq. (14), which decreases the effective magnitude of the excluded volume. It is worth noting that for more concentrated solutions the pairwise approximation ceases to become valid: from multi-particle theory Brady and Durlofsky (1988) obtained a value of 5 for hard spheres, also from a solvent frame of reference (Harding and Johnson 1985a, b).
On the grounds that the osmotic second virial coefficient equates with half of the thermodynamic contribution to excluded volume, the same situation also applies to its hydrodynamic counterpart. The excluded volume contribution to an experimentally measured concentration coefficient \({k}_{D}\) (mL/g) in Eq. (14) thus becomes [\(\left({\lambda }_{T}-{\lambda }_{H}\right){v}_{s}]\)/2 to be consistent with the description of \({\lambda }_{T}\) as \(2{B}_{22}^{HS}/{M}_{2}\).
Most of the subsequent attention has been directed towards the determination of D from dynamic light scattering studies, for which the same expression [Eq. (14)] has also been obtained for an uncharged spherical particle (Felderhof (1978; Wills 1979; Phillies and Wills 1981; Cichocki and Felderhof 1988). Petsev and Denkov (1992) have shown that the presence of net charge on those hard spheres increases the magnitude of the thermodynamic term in accordance with Eqs. (4a,4b) and (5). The corresponding relationship for the hydrodynamic term is dominated by the Oseen contribution)\(\left( {\lambda_{o} v_{s} } \right)\)
which establishes that the Oseen hard-sphere contribution to \({\lambda }_{H}\) is 6: the remainder (0.55) comes from two other contributions that account for short-range hydrodynamic interactions (Felderhof 1978; Petsev et al. 1992).
The coefficient describing the concentration dependence of the translational diffusion coefficient, \({k}_{D}\)(mL/g) = (\({\lambda }_{T}-{\lambda }_{H}){v}_{s}/2\), for charged hard spheres (HS) has therefore been considered to be given by the expression
with \({f}_{22}\left(r\right)\) defined by Eqs. (4b) and (5). Although the two integrals in Eq. (16) are usually evaluated by expanding the exponential in \({f}_{22}(r)\) as a power series in r, the value obtained for the electrostatic contribution to the excluded volume (\({\lambda }_{T}{v}_{s}\)) is an overestimate (Wills and Winzor 2009); and a similar situation presumably applies to the corresponding contribution to the hydrodynamic term \({\lambda }_{O}{v}_{s}\) in Eq. (15). We have therefore used the trapezoidal integration procedure for estimates of \(({\lambda }_{T}-{\lambda }_{H}){v}_{s}\) in considerations of experimental systems
In dynamic light scattering studies \(({\lambda }_{T}^{HS}-{\lambda }_{H}^{HS}\)) has often been taken as 1.45 for dilute solutions of rigid spherical particles in dominant Brownian motion, the value deduced by Batchelor (1976) for sedimentation velocity and traditional diffusion measurements, where particle flux is effected by a concentration gradient. It is worth pointing out that this value however only applies to very dilute conditions where the pairwise approximation is valid (Brady and Durlofsky 1988; Winzor et al 2021). Use of this value in dynamic light scattering studies has also been criticized by Phillies (1987) on the grounds that light scattering spectroscopy is merely sensitive to particle position; and that the value of \(({\lambda }_{T}^{HS}-{\lambda }_{H}^{HS}\)) should therefore be decreased to − 0.9.
Analysis of experimental results
Lysozyme represents a good starting point for analysis, due to its low degree of asymmetry (Blake et al 1965) and approximate uniform surface charge distribution (Fig. 1). The ionic strength dependence of the light scattering second virial coefficient for lysozyme in acetate and acetate–chloride buffers (pH 4.7) is presented in Fig. 2a, where the experimental points \(\left( \bullet \right)\) have been calculated from the values of \({A}_{2}\) reported in Table 2 of Muschol and Rosenberger (1995) and a molecular mass of 14,600 Da. Attempts to describe these data in thermodynamic terms, \({2A}_{2}{M}_{2}={\lambda }_{T}{v}_{s}-{\overline{v} }_{2}\) \(,\) with respective values of 1.9 nm and 11 for \({R}_{2}\) and \({Z}_{2}\) (Muschol and Rosenberger 1995) to solve the integrals in Eq. (4a,b) lead to their consistent overestimation \((- - -)\).

Electrostatic map calculated using the Poisson–Boltzmann equation of hen egg white lysozyme (PDB: 1AKI) at pH 7.0. It can be clearly seen that there is an even distribution of charge across the surface (positive: blue; negative: red). Hydrophobic patches are shown in grey: the only patch visible was c.a. 5 angstroms in diameter seen in the bottom right of the molecule, and is unlikely to contribute to aggregation due to the large electrostatic shadow cast by the other residues in the molecule. No other patches were visible on the molecule (data not shown) (color figure online)

Analysis of static light scattering data for lysozyme solutions at pH 4.7 (from data of Muschol and Rosenberger 1995). a Experimental results \(\left( \bullet \right)\) for the dependence of the second virial coefficient \({A}_{2}{M}_{2}\) upon ionic strength, together with theoretical dependence predicted either on the basis of its consideration as the equivalent of the osmotic second virial coefficient for protein self-interaction, \({\lambda }_{T}{v}_{s}/2\) \(\left( { - - - } \right)\), or the combination of that parameter and its hydrodynamic counterpart (—), as in Eq. (16) for the diffusion concentration dependence coefficient, \(\left({\lambda }_{T}-{\lambda }_{H}\right){v}_{s}/2\). b Demonstration of the correlation between \({k}_{D}\) and \({A}_{2}{M}_{2}\). [Data for \({\lambda }_{T}\) and \({\lambda }_{H}\) calculated from Table 2 of Muschol and Rosenberger (1995)]
This finding clearly corroborates earlier assertions that the light scattering coefficient should not be regarded as the osmotic second virial coefficient (Deszczynski et al 2006; Winzor et al. 2007; Wills et al. 2015). Much better agreement between experiment and prediction is achieved (_______) by adopting the viewpoint that \(2{A}_{2}{M}_{2}\) monitors [(\({\lambda }_{T}-{\lambda }_{H}){v}_{s}-{\overline{v} }_{2}]\) with the additional hydrodynamic term in \(\lambda_{H}\) (see also Harding and Johnson 1985a, b), an observation that favours consideration of the light scattering coefficient as a hydrodynamic rather than an equilibrium parameter. From Eq. (16) it is evident that the concentration dependence of \({A}_{2}{M}_{2}\) should then parallel that for the diffusion coefficient, which also monitors [\(({\lambda }_{T}-{\lambda }_{H}){v}_{s}/2]-{\overline{v} }_{2}\). The small extra term \({\overline{v} }_{2}\) comes from the Gibbs–Duhem relation (Harding & Johnson 1985a).
The extent of that correlation is shown in Fig. 2b, where the values of \({k}_{D}\) have also been calculated from Table 2 of Muschol and Rosenberger (1995) after correcting for their use of the unsolvated mole fraction (\({\overline{v} }_{2})\) for \({v}_{s}\). Although those results conform reasonably well with the concept of a linear relationship between \({k}_{D}\) and \({A}_{2}{M}_{2} \) with a slope of unity, the observation of a finite (negative) ordinate intercept precludes their consideration as the same parameter.
Inasmuch as Eq. (16) implies constancy of viscosity (taken as that of buffer, \(\eta_{b}\) ), no account has been taken of the effect of solution viscosity on the magnitude of the measured diffusion coefficient at finite protein concentrations. This deficiency is remedied by writing the concentration dependence of the diffusion coefficient as (Scott et al. 2014)
where η denotes the viscosity of a protein solution with concentration \({c}_{2}\) for which D was measured, and \(D_{o}\) that of buffer (the solution viscosity in the limit of zero solute concentration to which \({D}_{o}\) refers). Allowance for the fact that the relative viscosity is related to the intrinsic viscosity \(\left[ \eta \right]\) of a spherical protein species by the expression (Tanford 1961; Harding 1997)
introduces an additional decrease in the predicted value of \({k}_{D}\) by 2.5 \({v}_{S}\). An ordinate intercept of − 2.5 \({v}_{s}\) and a slope of unity is thus the predicted dependence of \({k}_{D}\) upon \({A}_{2}{M}_{2}\) (— in Fig. 2b). This is close to the intercept value of ~ − 3.1 \({v}_{s}.\) Exact agreement is made if allowance is made for the strong dependence of the Einstein-Simha viscosity shape factor ν on shape (ν = 2.5 for spheres and > 2.5 for other shapes), and can be determined exactly from triaxial crystallographic dimensions (Harding 1982, 1997; Harding et al 1979, 1981, 1982, 1983, 2005). From its crystallographic dimensions (Blake et al 1965) lysozyme approximates a prolate ellipsoid of axial ratio ~ 2.1. Using the program ELLIPS1 (Harding et al 2005; Garcia de la Torre & Harding 2013) this corresponds to a value of ν = 3.1. By contrast the exclusion volume term A2M2 is relatively insensitive to such a shape change (Rallison & Harding 1985; Harding et al 1999). The degree of conformity between experiment and prediction is considered excellent for the reported data even with no indication of experimental uncertainty inherent in the measurements.
Similar linear plots for the dependence of \(k_{D}\) upon \(A_{2} M_{2}\) have been reported for five monoclonal antibodies in histidine–chloride buffers (pH 6.0) with low and high ionic strengths (Lehermayr et al. 2011), and also for a single monoclonal antibody over a range of pH and ionic strengths (Roberts et al. 2014). Data from the former study are shown in Fig. 3a, and those at pH 5.0 and pH 5.75 from the latter investigation in Fig. 3b. A molar mass \({M}_{2}\) of 145 kDa and a Stokes radius \({R}_{2}\) of 5.2 nm (Roberts et al. 2014) have been used to calculate the predicted ordinate intercepts. As in Fig. 2b, the experimental results conform reasonably well with a slope of unity (— in Fig. 3a, b), although the intercepts are ~ 7 mL/g, considerably differ from the predicted − 2.5 \({v}_{S}\) for hard spheres. However, again if we allow for the greater sensitivity to shape of the viscosity increment ν compared with the exclusion volume: the value for the Einstein-Simha viscosity increment ν for an IgG antibody of molar mass ~ 150,000 g/mol is ~ 4.3 (Longman et al 2005). The time averaged hydration δ for antibodies is ~ 0.6 g water/g protein (Lu et al 2006) so \({v}_{S}\) = (\({\overline{v} }_{2}\)+ δ/ρo) ~ 1.35 mL/g, where ρo is the density of the aqueous solvent (~ 1 g/L). So the predicted value for the intercept = − ν\(.{v}_{S}\) is ~ 6–7 mL/g and appears to be consistent with Fig. 3a and b, and the experimentally measured values for the intrinsic viscosity (Kilar et al. 1985; Longman et al 2005).

Further evidence for correlation between the diffusion concentration dependence coefficient \({k}_{D}\) and the light scattering second virial coefficient \({A}_{2}{M}_{2}\) for monoclonal IgG antibodies. a Combined results for five monoclonal IgG antibodies (pH 6.0) at high and low ionic strengths. [Data taken from Lehermayr et al (2011)] b Corresponding dependence for a single monoclonal antibody at pH 5.0 and pH 5.75 and a range of ionic strengths. [Data from Roberts et al. (2014)]
Concluding remarks
The ionic strength dependence of \({A}_{2}{M}_{2}\) for lysozyme (pH 4.7) shown in Fig. 2a has again challenged the designation of this light scattering coefficient as the osmotic second virial coefficient, either in its molar form or its molal form \({C}_{22}{.M}_{2}={\lambda }_{T}{v}_{s}/2-{\overline{v} }_{2}\), which is unequivocally a thermodynamic parameter. Further support for the need to consider the Rayleigh ratio \(R_{\theta }\) obtained from “static” light scattering intensity as a steady-state rather than an equilibrium measurement comes from Figs. 2b, 3a b, which provide direct experimental evidence of a correlation between \({A}_{2}{M}_{2}\) and the corresponding parameter, \({k}_{D}\), for concentration dependence of the translational diffusion coefficient. This reflects the difference between the consequences of excluded volume and hydrodynamic intermolecular interactions. In other words, evidence is mounting for identification of the light scattering second virial coefficient as (\({\lambda }_{T}-{\lambda }_{H}){v}_{s}/2\) rather than \({\lambda }_{T}{v}_{s}/2\), and hence for cessation of its consideration as a thermodynamic parameter.
By demonstrating that the light scattering second virial coefficient \({A}_{2}{M}_{2}\) equates with the excluded volume contribution to \({k}_{D}\), the coefficient describing the concentraton dependence of diffusion coefficients obtained by dynamic light scattering, this investigation has substantiated our earlier conclusion (Deszczynski et al. 2006; Winzor et al. 2007) that \({A}_{2}\) should not be designated as the osmotic second virial coefficient for protein self-interaction, \(B_{22} /M_{2}^{2}\) – a parameter with thermodynamic status (McMillan and Mayer 1945). A subsequent challenge to that conclusion (Blanco et al. 2011) perhaps reflected an entrenched historical practice whereby the nonideality parameter emanating from scattering spectroscopy measurements (visible light, X-rays, neutrons) is automatically described as the osmotic second virial coefficient. Indeed, the possibility that negative values of \({A}_{2}\) might include some consequences of hydrodynamic interactions had already been suggested (Neal et al. 1999). That suggestion appears to be further supported by the demonstration that \({A}_{2}{M}_{2}\) monitors (\({\lambda }_{T}-{\lambda }_{H}\)), the difference between thermodynamic and hydrodynamic excluded volume contributions, respectively; and thereby appears to invalidate the implication inherent in the light scattering literature that \({A}_{2}\) is the thermodynamic parameter \({B}_{2}\) (= B22/M 2) in mL.mol/g2. Specifically
A second problem addressed in this investigation has been the failure of the dependence of \({k}_{D}\) upon \({A}_{2}{M}_{2}\) to pass through the origin despite the 1:1 correlation. That dilemma has been overcome by incorporating the additional effect of solution viscosity on D–c dependence, a factor neglected by Batchelor (1976) and Felderhof (1978) in the rationalization of nonideality on the statistical-mechanical basis of excluded volume.
Finally, some consideration needs to be given to the consequences of the current reclassification of \({A}_{2}\) as a non-thermodynamic parameter on recent procedures for the quantitative characterization of nonideality in static light scattering measurements (Minton 2007; Fernàndez and Minton 2009). From the theoretical expression that forms the basis of the analysis for a single uncharged solute,
it is evident that Rayleigh scattering ratio \(R_{\theta }\) is being accorded full thermodynamic status: \(8{v}_{s}\) is \(\lambda_{T}\) for an uncharged sphere in the above considerations. Incorporation of the increased excluded volume for a charged species, \(\lambda_{T}^{EL} ,\) is accommodated (Minton and Edelhoch 1982) by increasing the effective size of the hard sphere by expressing Eq. (7) as
The method is empirical in the sense that a value for the solvated specific volume is obtained as a curve-fiting parameter, whereupon the contributions of the hydrodynamic terms \(\lambda_{H}^{HS} ,\,\lambda_{H}^{EL}\) are also incorporated. It therefore remains valid as an empirical procedure despite being based on thermodynamic expressions that do not apply to static light scattering measurements. There is also potential for error in the analysis arising from the other curve-fitting parameter, which has been taken as \({M}_{2}\) in Eq. (19) but which incorporates a protein–cosolute virial coefficient term \({B}_{23}{C}_{3}\) because of the need to regard buffer species as additional non-scattering cosolutes (Kirkwood and Goldberg 1950; Winzor et al. 2007; Blanco et al. 2011; Wills and Winzor 2017).
The combination of empiricism and the non-equilibrium nature of \(A_{2}\) clearly detracts from “static” light scattering as an alternative to techniques such as osmometry and sedimentation equilibrium, where conformity with the constraints of constant temperature and solvent chemical potential fully justify the interpretation of experimental data in terms of thermodynamic expressions for single-solute systems. This can also have a bearing on the analysis of reversible self-association processes. With sedimentation equilibrium (and osmotic pressure the effects of reversible self-association are nonetheless difficult to extract from thermodynamic nonideality effects as they are both dependent on concentration (see Scott and Winzor 2009 and Harding and Rowe 2010) and sometimes these effects can cancel out to lead to “pseudo-ideal” conditions. Where structural information about the monomer species is available, for globular based structures the best procedure is to calculate B2 or B22 from the tri-axial shape of the molecule (and the net charge and ionic strength) using a routine COVOL, downloadable from https://www.nottingham.ac.uk/ncmh/ (Harding et al 1998, 1999; Harding 2013)—or for asymmetric shapes such as immunoglobulins using SOLPRO (Garcia de la Torre et al. 1999). With light scattering there may be an extra layer of difficulty with having the added complication of hydrodynamic as well as thermodynamic nonideality to deal with.
References
Batchelor GK (1976) Brownian diffusion of particles with hydrodynamic interaction. J Fluid Mech 74:1–29
Behlke J, Ristau O (1999) Analysis of the thermodynamic nonideality of proteins by sedimentation equilibrium experiments. Biophys Chem 76:13–23
Blake C, Koenig D, Mair G, North ACT, Phillips DC (1965) Sarma VR (1965) Structure of hen egg-white lysozyme: a three-dimensional Fourier synthesis at 2 Å resolution. Nature 206:757–761. https://doi.org/10.1038/206757a0
Blanco M, Sahin E, Li Y, Roberts CJ (2011) Reexamining protein–protein and protein–solvent interactions from Kirkwood-Buff analysis of light scattering in multicomponent systems. J Chem Phys 134(22):25103
Brady JF, Durlofsky J (1988) The sedimentation rate of disordered suspensions. J Phys Fluids 31:717–727
Cantor CR, Schimmel PR (1980) Biophysical Chemistry, Freeman, San Francisco, CA. Chapter 10–3
Cichocki B, Felderhof BU (1988) Short-term diffusion coefficients and high-frequency viscosity of dilute suspensions of spherical Brownian particles. J Chem Phys 89:1039–1064
Deszczynski M, Harding SE, Winzor DJ (2006) Second virial coefficients as predictors of protein growth: evidence from sedimentation equilibrium studies that refutes the designation of those light scattering parameters as osmotic second virial coefficients. Biophys Chem 120:106–113
Felderhof BU (1978) Diffusion of interacting Brownian particles. J Phys a: Math Gen 11:929–937
Fernàndez C, Minton AP (2009) Static light scattering from concentrated protein solutions. II. Experimental test of theory for protein mixtures and weakly self-associating proteins. J Chem Phys 96:1992–1998
García de la Torre J, Harding SE (2013) Hydrodynamic modelling of protein conformation in solution: ELLIPS and HYDRO. Biophys Rev 5:195–206
García de la Torre J, Harding SE, Carrasco B (1999) Calculation of NMR relaxation, covolume, and scattering-related properties of bead models using the SOLPRO computer program. Europ Biophys J 28:119–132
George A, Wilson WW (1994) Predicting protein crystallization from a dilute solution property. Acta Cryst D50:361–365
Guo B, Kao S, McDonald H, Asanov A, Coombs LL, Wilson WW (1999) Correlation of second virial coefficients and solubilities useful in protein crystal growth. J Cryst Growth 196:424–431
Hall DR, Harding SE, Winzor DJ (1999) The correct analysis of low-speed sedimentation equilibrium distributions recorded by the Rayleigh interference optical system in a Beckman XL-I ultracentrifuge. Prog Coll Polym Sci 113:62–68
Harding SE (1982) A computer program for evaluating the hydrodynamic parameters for a macromolecule for any given value of its axial dimensions. Comput Biol Med 12:75–80
Harding SE (1985) The representation of equilibrium solute distributions for non-ideal polydisperse systems in the analytical ultracentrifuge. Biophys J 47:247–250
Harding SE (1997) The intrinsic viscosity of biological macromolecules. Progress in measurement, interpretation and application to structure in dilute solution. Prog Biophys Mol Biol 68:207–262
Harding SE, Cölfen H, Aziz Z (2005) The ELLIPS suite of whole-body protein conformation algorithms for Microsoft Windows. In: Scott DJ, Harding SE, Rowe AJ (eds) Analytical Ultracentrifugation. Techniques and Methods. Royal Society of Chemistry, Cambridge, UK, pp 460–483
Harding SE, Dampier MJ, Rowe AJ (1979) Modelling biological macromolecules in solution: the general triaxial ellipsoid can now be employed IRCS. Med Sci 7:33
Harding SE, Dampier MJ, Rowe AJ (1981) The viscosity increment of a dispersion of triaxial ellipsoids under dominant brownian motion. J Coll Int Sci 79:7–13
Harding SE, Dampier MJ, Rowe AJ (1982) The viscosity increment for ellipsoids of revolution: some observations on the Simha formula. Biophys Chem 15:205–208
Harding SE, Horton JC, Jones S, Thornton JM, Winzor DJ (1999) COVOL: An interactive program for evaluating second virial coefficients from the triaxial shape or dimensions of rigid macromolecules. Biophys J 76:2432–2438
Harding SE, Horton JC, Winzor DJ (1998) COVOL: an answer to your thermodynamic non-ideality problems? Biochem Soc Trans 26:737–740
Harding SE, Johnson P (1985a) On the concentration dependence of macromolecular parameters. Biochem J 231:543–547
Harding SE, Johnson P (1985b) Physicochemical studies of turnip-yellow-mosaic virus: homogeneity, relative molecular masses, hydrodynamic radii and concentration dependence of parameters in non-dissociating solvents. Biochem J 231:549–555
Harding SE, Rowe AJ (2010) Insight into protein-protein interactions from analytical ultracentrifugation. Biochem Soc Trans 38:901–907
Harding SE (2013) ELLIPS and COVOL. In: Roberts, G.C.K, ed., Encyclopedia of Biophysics, Springer Verlag, Berlin. Pp 654–662
Hill TL (1959) Theory of solutions. 2. osmotic virial expansion and light scattering in two component solutions. J Chem Phys 30:93–97
Hill TL (1968) Thermodynamics for Chemists and Biologists. Addison-Wesley, Reading, MA
Van Holde KE (1985) Physical Biochemistry. Prentice-Hall, Englewood Cliffs, NJ
Kilar F, Simon I, Lakatos S, Vonderviszt F, Medgyesi GA, Zavodszky P (1965) Conformation of human IgG subclasses in solution. small-angle x-ray scattering and hydrodynamic studies. Eur J Biochem 147:17–25
Kirkwood JG, Goldberg RJ (1950) Light scattering arising from composition fluctuations in multicomponent systems. J Chem Phys 18:54–57
Lehermayr C, Mahler H-C, Mäder K, Fischer S (2011) Assessment of net charge and protein–protein interactions of different monoclonal antibodies. J Pharm Sci 100:2441–3256
Longman E, Harding SE, Marheineke N (2005) Identifying differences in solution conformation of two chimeric IgG3 antibodies through triple detection SEC LCGC (Liquid Chromatography, Gas Chromatography) 18: 662–668
Lu Y, Longman E, Davis KG, Ortega A, Grossmann JG, Michaelsen TE, Garcia de la Torre J, Harding SE (2006) Crystallohydrodynamics of protein assemblies: combining sedimentation, viscometry, and x-ray scattering. Biophys J 91(5):1688–1697
Mayer JE (1950) Theory of ionic solutions. J Chem Phys 18:1426–1436
McMillan WG, Mayer JE (1945) The statistical thermodynamics of multicomponent systems. J Chem Phys 18:276–305
Minton AP (2007) Static light scattering from concentrated protein solutions. I. general theory for protein mixtures and application to self-associating proteins. Biophys J 93:1321–1328
Minton AP, Edelhoch H (1982) Light scattering of bovine serum albumin solutions: extension of the hard particle model to allow for electrostatic repulsion. Biopolymers 21:451–458
Mos HJ, Pathmamanoharan C, Dhont JKG, de Kruif CG (1986) Scattered light intensity cross correlation. II Experiments J Chem Phys 84:45–49
Muschol M, Rosenberger F (1995) Interactions in undersaturated and supersaturated lysozyme solutions: static and dynamic light scattering results. J Chem Phys 103:10424–10432
Neal BL, Asthagiri D, Velev OD, Lenhoff AM, Kaler EW (1999) Why is the osmotic second virial coefficient related to protein crystallization? J Cryst Growth 196:377–387
Petsev DN, Denkov ND (1992) Diffusion of charged colloidal particles at low volume fraction: theoretical model and light scattering experiments. J Coll Interface Sci 149:329–344
Phillies GJD (1987) Numerical interpretation of the concentration dependence of micelle diffusion coefficients. J Coll Interface Sci 119:518–523
Phillies GJD, Wills PR (1981) Light scattering spectrum of a suspension of interacting Brownian macromolecules. J Chem Phys 75:508–514
Rallison JM, Harding SE (1985) Excluded volumes for pairs of triaxial ellipsoids at dominant Brownian motion. J Coll Int Sci 103:284–289
Roberts D, Keeling R, Tracka M, van der Walle CF, Udin S, Warwicker J, Curtis R (2014) The role of electrostatics in protein–protein interactions of a monoclonal antibody. Mol Pharmaceutics 11:2475–2489
Rosenbaum DF, Zukoski CF (1996) Protein interactions and crystalliation. J Cryst Growth 169:752–758
Scott DJ, Harding SE, Winzor DJ (2014) Concentration dependence of translational diffusion coefficients for globular proteins. Analyst 139:6242–6248
Scott DJ, Winzor DJ (2009) Comparison of methods for characterizing non-ideal self-association by sedimentation equilibrium. Biophys J 97:886–896
Serdyuk IN, Zaccai NR, Zaccai J (2007) Methods in Molecular Biophysics, Cambridge University Press, Parts C & D.
Stockmayer WH (1950) Light scattering in multicomponent systems. J Chem Phys 18:58–61
Tanford C (1961) Physical Chemistry of Macromolecules. Wiley, New York
Vivarés D, Bonneté F (2002) X-ray scattering studies of Aspergillus flavus urate oxidase: towards a better understanding of PEG effects on the crystallization of large proteins. Acta Cryst D58:472–479
van der Waals JD (1873) Over de Continuiteit van den Vloeistofroestand. Leiden University, The Netherlands
Wills PR (1979) Isothermal diffusion and quasielastic light scattering of macromolecular solutes at finite concentrations. J Chem Phys 70:5865–5874
Wills PR, Comper WD, Winzor DJ (1993) Thermodynamic nonideality in macromolecular solutions: interpretation of virial coefficients. Arch Biochem Biophys 300:206–212
Wills PR, Scott DJ, Winzor DJ (2015) The osmotic second virial coefficient for protein self-interaction: use and misuse to describe thermodynamic nonideality. Anal Biochem 490:55–65
Wills PR, Winzor DJ (2005) van der Waals transition in protein solutions. Acta Cryst D61:832–836
Wills PR, Winzor DJ (2009) Direct allowance for effects of thermodynamic nonideality in the quantitative characterization of protein self-association by osmometry. Biophys Chem 145:64–71
Wills PR, Winzor DJ (2017) Rigorous analysis of static light scattering measurements on buffered protein solutions. Biophys Chem 228:108–113
Wills PR, Winzor DJ (1992) Thermodynamic nonideality and sedimentation equilibrium. In: Harding SE, Rowe AJ, Horton J (eds) Analytical Ultracentrifugation in Biochemistry and Polymer Science. Royal Society of Chemistry, Cambridge UK, pp 311–330
Winzor DJ, Carrington LE, Harding SE (2001) Analysis of thermodynamic non-ideality in terms of protein solvation. Biophys Chem 93:231–240
Winzor DJ, Deszczynski M, Harding SE, Wills PR (2007) Nonequivalence of second virial coefficients from sedimentation equilibrium and static light scattering studies of protein solutions. Biophys Chem 128:46–55
Winzor DJ, Dinu V, Scott DJ, Harding SE (2021) Quantifying the concentration dependence of sedimentation coefficients for globular proteins: a continuing age-old problem. Biophys Rev 13:273–288
Winzor DJ, Wills PR (1994) Thermodynamic nonideality and protein solvation. In: Gregory RB (ed) Protein-Solvent Interactions. Marcel Dekker, New York, pp 483–520
Acknowledgements
DJS was partially supported in the preparation of this manuscript by ISIS spallation neutron and muon source (UK). SEH was supported by the UK Biotechnology and Biological Sciences Research Council [grant number BB/T006404/1] and the Norwegian Ministry of Education and Research.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
“Special Issue: Analytical Ultracentrifugation 2022”.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Winzor, D.J., Dinu, V., Scott, D.J. et al. Experimental support for reclassification of the light scattering second virial coefficient from macromolecular solutions as a hydrodynamic parameter. Eur Biophys J 52, 343–352 (2023). https://doi.org/10.1007/s00249-023-01665-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00249-023-01665-w