

Regulation of CRISPR-Associated Genes by Rv1776c (CasR) in Mycobacterium tuberculosis
by
 ,
,
and
,
,
and
Wenping Wei
1,
Xiaofang Jiang
2,
Li Zhang
2,
Yunjun Yan
1,*,
Jinyong Yan
1,
Li Xu
1,

Chun-Hui Gao
3 and
Min Yang
1,*
1
Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan 430074, China
2
State Key Laboratory of Agricultural Microbiology, College of Life Science and Technology, Huazhong Agricultural University, Wuhan 430070, China
3
State Key Laboratory of Agricultural Microbiology, College of Resources and Environment, Huazhong Agricultural University, Wuhan 430070, China
*
Authors to whom correspondence should be addressed.
Biomolecules 2023, 13(2), 400; https://doi.org/10.3390/biom13020400
Submission received: 29 November 2022 / Revised: 7 February 2023 / Accepted: 18 February 2023 / Published: 20 February 2023
(This article belongs to the Special Issue Virulence Factors in Mycobacterium tuberculosis Infection: Structural and Functional Studies)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:
The CRISPR-Cas system is an adaptive immune system for many bacteria and archaea to defend against foreign nucleic acid invasion, and this system is conserved in the genome of M. tuberculosis (Mtb). Although the CRISPR-Cas system-mediated immune defense mechanism has been revealed in Mtb, the regulation of cas gene expression is poorly understood. In this study, we identified a transcription factor, CasR (CRISPR-associated protein repressor, encoded by Rv1776c), and it could bind to the upstream DNA sequence of the CRISPR-Cas gene cluster and regulate the expression of cas genes. EMSA and ChIP assays confirmed that CasR could interact with the upstream sequence of the csm6 promoter, both in vivo and in vitro. Furthermore, DNA footprinting assay revealed that CasR recognized a 20 bp palindromic sequence motif and negatively regulated the expression of csm6. In conclusion, our research elucidates the regulatory effect of CasR on the expression of CRISPR-associated genes in mycobacteria, thus providing insight into gene expression regulation of the CRISPR-Cas system.
1. Introduction
Clustered regularly interspaced short palindromic repeats (CRISPR) and CRISPR-associated (Cas) proteins are found in most (~90%) sequenced archaeal genomes and many (~50%) sequenced bacterial genomes [1,2]. In addition to its role as a prokaryotic adaptive immune system in protecting cells from the invasion of foreign mobile genetic elements, including bacteriophages and plasmids, the CRISPR-Cas system also modulates multiple biological processes such as gene regulation and bacterial virulence, especially pathogen stress responses [3,4,5]. However, the regulatory mechanism of the CRISPR-Cas system is far from clear.
Based on different modes of action and the function of Cas proteins, CRISPR-Cas systems are divided into two classes and six types. The Class 1 CRISPR-Cas system, which includes types I, III, and IV, can form multi-subunit complexes around the Cas7 backbone, whereas, in Class 2 (types II, V, and VI), interference is achieved by a single effector protein comprising multiple nuclease domains [1,5]. CRISPR-Cas systems immunity consists of three distinct immune stages: adaptation, RNA maturation, and interference [6]. The adaptation process involves the insertion of short DNA sequences called spacers, which originated from invading bacteriophages, into the CRISPR arrays to preserve the memory of previous infections. CRISPR arrays are transcribed and processed into CRISPR RNAs (crRNAs) that contain individual spacer sequences and are arranged by repeat fragments [7,8]. During the CRISPR interference stage, Cas proteins bind to individual crRNAs to form effector complexes that recognize complementary sequences (known as ‘protospacer’) in foreign nucleic acids. Once a protospacer is recognized, the target DNA or RNA transcript is degraded by a Cas-crRNA ribonucleoprotein complex [9,10,11].
Mycobacterium tuberculosis (Mtb), the causative agent of tuberculosis, encodes a complete type III-A CRISPR-Cas system in its genome [12]. The type III CRISPR-Cas system is one of the most refined, powerful, and versatile characterized CRISPR interference complexes. This system comprises Csm1/2/3/4/5/6, and Cas6, which are responsible for crRNA processing and maturation, and two adaptation proteins (Cas1 and Cas2) located next to two CRISPR arrays with 24 and 18 repeats, respectively [13,14,15]. The type III CRISPR-Cas system in M. bovis is identical to that in Mtb, except that the csm6 gene is truncated, and that spacer leader sequence is lacking [12]. A recent study has reported that the CRISPR-Cas system can effectively reduce plasmid transformation [16]. Research on the reconstruction and characterization of the Mtb CRISPR system in Escherichia coli has indicated that mycobacterial immunity to foreign nucleic acids is achieved mainly by cA6-activated Csm6 nuclease activity rather than through DNA cleavage by Csm1 [17]. Therefore, the regulation of csm6 gene expression is crucial for the biological activity of the CRISPR-Cas system in Mtb.
The genome of Staphylococcus epidermidis also harbors a type III A-CRISPR-Cas system, in which Csm6 is required for defense against the invasion of bacteriophages; Csm6 can degrade RNA transcripts to ensure efficient immunity under the conditions of slow degradation of target DNA [18]. This system provides effective immunity, but it may also lead to the growth arrest of infected cells during the interference stage, thus necessitating ribonuclease inactivation in the case of bacteriophage clearance [19,20]. Therefore, it is vital for bacteria to precisely regulate the expression of CRISPR-associated genes. In some bacterial species, transcription factors such as CPR (cyclic AMP receptor protein), LRP (leucine-responsive regulatory protein), LeuO (regulator of leucine biosynthesis operon), and H-NS (histone-like nucleoid-structuring protein) regulate the expression of the CRISPR-Cas system by binding to CRISPR promoters [21]. In Mtb, Rv2837c (CnpB) controls the expression of the CRISPR-Cas system through a nanoRNA with oligoribonuclease (Orn)-like activity [22]. However, there are few reports on transcription factors regulating the CRISPR-Cas system in mycobacteria.
In the present study, we identified a TetR family transcription factor and named it CasR; it regulated the expression of cas genes of the CRISPR-Cas system in Mtb. In vitro EMSA and in vivo ChIP assays showed that CasR specifically bound to the promoter of csm6 in the CRISPR-Cas system. The β-galactosidase activity assay and qPCR assay indicated that CasR negatively regulated the expression of csm6. We found that the CasR regulator was conserved in multiple mycobacteria, suggesting that it may control the CRISPR-Cas system broadly. Our findings deepen the understanding of gene regulation of the bacterial CRISPR-Cas system.
2. Materials and Methods
2.1. Bacterial Strains, Plasmids, and Reagents
Mtb H37Ra (purchased from ATCC) was cultivated in Middlebrook 7H9 medium containing 0.05% glycerol and 10% oleic acid–albumin–dextrose–catalase (OADC) enrichment. Escherichia coli BL21 (DE3) was incubated in Luria-Bertani (LB) medium and used for protein expression. The restriction enzymes, T4 DNA Ligase, Taq DNA polymerase, and DNase I were purchased from New England Biolabs (Beverly, MA, USA) and Thermo Fisher Scientific (Waltham, MA, USA). All antibiotics, including kanamycin (Kan) and ampicillin (Amp), were purchased from Sigma-Aldrich (Darmstadt, Germany). Antiserum was obtained from Dia-An Biotechnology Co., Ltd., (Wuhan, China). All constructed plasmids and strains are listed in Table S1.
2.2. Plasmid Construction and Protein Purification
To construct the casR-overexpressing strain, the casR coding sequence was amplified via PCR using a pair of respective primers (Table S2). The PCR products were digested with EcoRI/XbaI and then cloned into the EcoRI/XbaI-digested pMV261 vector with L5 regulatory promoter using T4 DNA ligase. The pMV261-casR recombinant plasmid was electroporated into M. tuberculosis H37Ra to generate an overexpression strain. For CasR and other regulator proteins’ purification, the PCR products were inserted into the pET28a vector of EcoRI/XbaI digestion using recombinant enzymes (Yeasen, Wuhan, China). Then, the recombinant plasmids were transformed into BL21 competent cells, and transformants were selected on plates containing 50 μg/mL Kan. Then, the cultures were grown at 37 °C until the OD600 (optical density) reached ~ 0.6 and were induced by adding 0.3 mM isopropyl β-D-thiogalactoside (IPTG, CAS 367-93-1, Amresco, Solon, OH, USA) for 16 h at 16 °C. The recombinant proteins were purified using an Ni2+-affinity column, as described previously [23]. The purified proteins were analyzed using SDS-PAGE. The supernatant was filtered with a 0.22 μm membrane and was concentrated by centrifugation at 2000× g at 4 °C using an ultrafiltration tube (molecular weight cut-off: 3 kDa, Millipore). Finally, the proteins were dialyzed using dialysis buffer (20 mM Tris-HCl, 100 mM NaCl, 10% glycerin, pH 8.0) for 2 h at 4 °C and stored at −80 °C. The protein concentration was determined using the Coomassie brilliant blue protein kit (Sigma-Aldrich, St. Louis, MO, USA).
2.3. Electrophoretic Mobility Shift Assay (EMSA)
The DNA probes used in EMSA were 500 base pairs (bp) long, including 400 bp upstream from the start codon and the first 100 bp of each gene coding region. All DNA probes were obtained via PCR using Mtb H37Ra genomic DNA as a template. The promoter primers for Rv0324, Rv1255c, Rv1473A, csm6, casR, and cas6 are listed in Table S2. DNA–protein interactions were performed as previously described, with some modifications [24]. Briefly, 20 μL of reaction mixtures containing 100 ng DNA probes, 0.5–2 μM regulator proteins and reaction buffer (100 mM Tris-HCl, pH 7.5, 7 mM MgCl2, 50 mM NaCl, 10% glycerin) were incubated for 30 min at room temperature. Then, the mixtures were electrophoresed into a 5% native polyacrylamide gel containing 0.5 × TBE buffer and ran at 150 V for 1 h. Images were acquired with a Typhoon Scanner (GE healthcare, Buckinghamshire, UK).
2.4. Chromatin Immunoprecipitation (ChIP) Assay
In vivo interactions of the CasR protein with its own promoter and potential target gene promoters were analyzed using the ChIP assay, using a previously reported method with some modifications [24]. In order to obtain a high-affinity CasR antibody, the high-purity CasR protein was sent to Dia-An Biotechnology Co., Ltd. (Wuhan, China) for polyclonal antibody production. The Mtb H37Ra strain was cultured in 100 mL of 7H9 medium supplemented with 10% OADC until OD600 reached 1.0. The bacterial solution was fixed with 1% formaldehyde for 30 min at room temperature, then 0.125 M glycine was added to terminate the reaction. Afterwards, cross-linked cells were harvested, washed with 5 mL of pre-cooled PBS, and resuspended in 1 mL of TBSTT-I (20 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween 20, 0.1% Triton X-100, 1 mM protease inhibitor cocktail). The sample was sonicated at 250 W on ice for 3 min, and the average 0.5–1 kb DNA fragment was selected for further experiments. A 100 μL sample of the supernatant was used as the input fraction, whereas the remaining 800 μL was divided into two parts. These two parts were added with preimmunization serum or antiserum against CasR and incubated under rotation for 3 h at 4 °C. The protein complexes were immunoprecipitated with 50 μL of 50% protein A agarose under rotation for 8 h at 4 °C. The immune complexes were washed 5 times with 1 mL of TBSTT-I, 1 time with TBSTT-II (500 mM NaCl), and 1 time with TBSTT-III (containing 0.5% Tween 20 and 0.5% Triton X-100). Finally, the complexes were dissociated with 50 μL of TE (containing 20 mM Tris-HCl and 10 mM EDTA, 0.5% SDS, pH 7.8) in a 65 °C water bath for 2 h. The DNA samples from the ChIP assay were purified and analyzed using PCR. The primer sequences were listed in Table S2. The PCR amplification was performed as follows: pre-denaturation at 95 °C for 5 min, followed by 25 cycles of 95 °C for 20 s, 60 °C for 20 s, and 72 °C for 30 s. The PCR products were separated using 1.5% agarose gel electrophoresis.
2.5. DNase I Footprinting Assay
For the footprinting assay, the 500 bp casRp DNA fragment at the positions from −400 to +100 of the casR translational start site was amplified via PCR using specific primers labeled with 5′-FAM. A DNase I footprinting assay was performed, as described previously [25]. The amplified DNA products were purified with the OMEGA DNA Purification kit (OMEGA), and then subjected to the binding reaction under the same conditions as EMSA. Briefly, a total of 1.5 μg of DNA probes were co-incubated with CasR protein in reaction buffer for 30 min at 25 °C. Then, 0.0025 U DNase I was added to the mixture for digestion at 37 °C for 100 s, and phenol chloroform was added immediately to terminate the reaction. The samples were centrifuged and then precipitated with ethanol at −80 °C overnight. Finally, the DNA precipitates were resuspended in 10 μL of ddH2O, and the fragments were subjected to STR sequencing by Tsingke Biological Technology (Wuhan, China). Electropherograms were analyzed using GENEMAPPER software (version 4.0, Applied Biosystems).
2.6. β-Galactosidase Activity Assay
A series of operon-lacZ fusion plasmids derived from pMV261 were constructed to perform β-gal activity experiments in the M. smegmatis strain [26]. The target and control gene promoter fragments were amplified with PCR using two pairs of primers (Table S2), and then cloned into the pMV261-lacZ backbone. Similarly, the regulator casR was ligated behind the target promoter and cloned into the pMV261-lacZ vector together. To construct the csm6pm-casR-lacZ or casRpm-casR-lacZ plasmid, which were used as a control for promoter mutations, the mutant promoter csm6pm or casRpm was inserted upstream of the casR gene. In addition, the enhanced promoter hsp60 was inserted into pMV261-lacZ as a positive control, and the empty plasmid served as a negative control. All the recombinant plasmids were transformed into the M. smegmatis strain to obtain corresponding reporter strains. All the reported strains were verified by PCR, using the upstream pMV261 sequence of the 5′-end of the DNA fragment as a forward primer and the 3′-end of the DNA fragment as the reverse primer. Then, the PCR products were sent to the TsingKe Company for DNA sequencing. The strains were cultured in 5 mL of 7H9 medium until the mid-log phase. The cells were harvested and washed using cold PBS. β-gal activity was measured as described previously [27].
2.7. Real-Time Quantitative Reverse-Transcription PCR (qRT-PCR) Assay
The pMV261 empty and casR-overexpressing strains were grown to OD600 of 1.0 in 100 mL of 7H9 medium. The extraction of mRNA from the cultures and qRT-PCR analysis was performed as described previously [23]. The cDNA was obtained using the HiScript II Q RT kit (Vazyme, Nanjing, China). The qRT-PCR system consisted of 20 μL of solution containing 10 μL of 2 × SYBR Green qPCR mix, 400 nM specific primers, and 1 μL of cDNA. The reactions were performed in a Bio-Rad CFX instrument under the following program: 95 °C for 1 min and 40 cycles of 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 30 s. The expression level of each gene was normalized with sigA as an internal reference. Gene relative expression was determined according to the 2−ΔΔCt method [28].
3. Results
3.1. Identification of Transcription Factors Regulating CRISPR-Cas System Gene Expressions
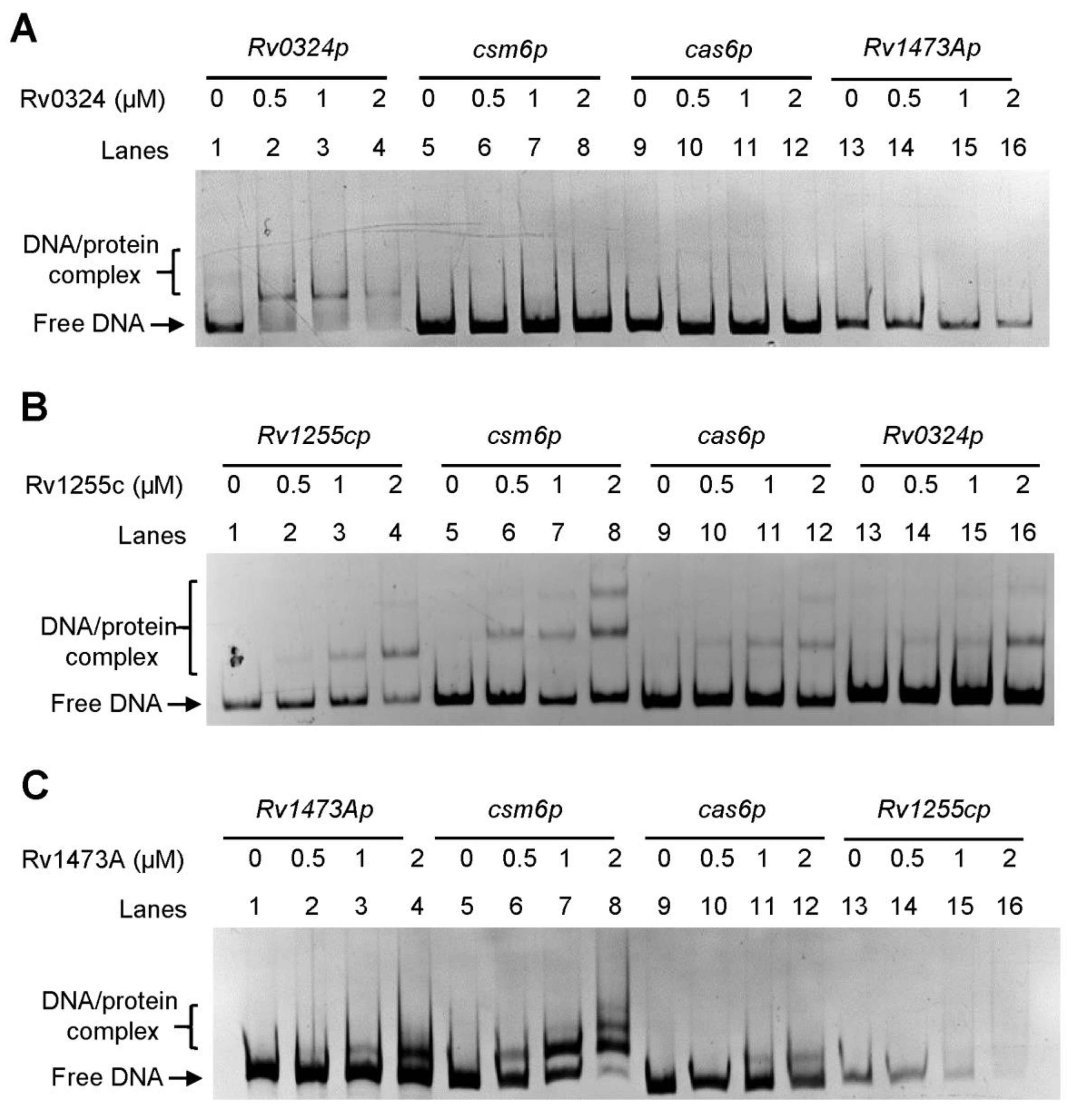
In M. tuberculosis, the CRISPR-Cas system is a type III-A system. The expression pattern of csm6 may differ from that of other cas genes, and the transcription start site (TSS) has been identified as an independent one upstream of the csm6 by high-throughput screening (Figure 1A) [29]. To find putative regulators of the CRISPR-Cas system, we queried the TB database [30,31] and retrieved seven regulators encoded by Rv0047c, Rv0324, Rv1255c, Rv1473A, Rv1776c, Rv2034, and Rv3133c (devR), which may interact with the gene promoter region of csm6 (Figure 1B). Among these regulators, Rv2034 and Rv3133c were both global ones which can regulate multiple biological processes, including hypoxia adaptation, and virulence; moreover, these two transcriptional factors have been extensively investigated [32,33]. However, knowledge of the other transcriptional factors was still lacking. Notably, the ChIP-seq signal of Rv0324 and Rv1473A was 10-fold and 2-fold more intensive than that of Rv2034 and Rv3133c, respectively [31]. In addition, evidence showed that the binding regions of Rv1473A, Rv1255c, and Rv0047c are located upstream of the csm6 gene, while the binding regions of Rv1776c and Rv0324 are located in the coding region of the csm6 gene. Based on these findings, we hypothesized that Rv0047c, Rv0324, Rv1255c, Rv1473A, and Rv1776c might be novel regulators of the CRISPR–Cas system in M. tuberculosis. To verify our hypothesis, we subsequently cloned, expressed, and purified these regulators. All of these regulators were expressed successfully, except that Rv0047c was completely insoluble and failed to result in purified protein (Figure S1). Furthermore, the EMSA was performed to determine whether the remaining regulators (excluding Rv0047c) could specifically interact with two putative promoters of csm6 (csm6p) and cas6 (cas6p). As shown in Figure 2A, Rv0324 formed a protein–DNA complex shift band with its own promoter, but it did not bind to csm6p, cas6p, or the promoter region of Rv1473A (Rv1473Ap). Moreover, Rv1255c bound to self-promoter, csm6p, cas6p, Rv0324p, and some unrelated promoters, indicating that Rv1255c lacked specific binding activity to DNA (Figure 2B and Figure S2). Similar to Rv1255c, Rv1473A also lacked specific DNA binding activity, except for the different position where its shift band appeared (Figure 2C and Figure S3). After the initial filtration, Rv1776c was screened as the only candidate and named CasR.
3.2. CasR Specifically Interacts with csm6 Promoter In Vivo and In Vitro
We then analyzed the interaction of CasR protein with several DNA probes, whose sizes and characteristics are given in Figure 3A. Notably, when either casRp or csm6p DNA probes were co-incubated with increasing concentrations of CasR (0, 0.5, 1, and 2 μM), clearly shift bands were observed (Figure 3B, Lanes 1–8). In contrast, CasR did not bind to cas6p or the control promoter groEL1p (Figure 3B, Lanes 13–16 and Figure S4). These results suggested that CasR could specifically bind to its own promoter region and the promoter of csm6 in vitro.
A ChIP assay was performed to further confirm the binding of CasR to casRp and csm6p DNA fragments in M. tuberculosis. As shown in Figure 3B, after immunoprecipitation with the CasR antiserum, two DNA fragments were specifically recovered, but they were not recovered after immunoprecipitation with the pre-immune serum (Figure 3C). In contrast, the promoter of cas6 (as negative control) was not recovered by either CasR antiserum or pre-immune serum (Figure 3C). These results showed that CasR specifically interacted with the upstream DNA sequence of csm6, both in vitro and in vivo.
Orthologs of CasR were identified based on sequence similarity and the conservation of adjacent genes, and CasR and its orthologs were annotated as hypothetical proteins (Figure 3D). The CasR region was highly conserved in M. tuberculosis H37Rv, M. tuberculosis H37Ra, and M. bovis BCG with amino acid identity of 100% but not highly conserved in M. smegmatis MC2 155, with an amino acid identity of only 59.55% (Figure S5). The NCBI database showed that the gene casR was 561 bp long, and the protein encoded 186 amino acids containing a typical TetR_N superfamily domain with a helix-turn-helix DNA binding motif, and a molecular weight of 20314 Daltons (Figure 3E), suggesting that CasR belongs to the TetR family transcription factors.
3.3. CasR Recognizes A 20 bp Palindrome Sequence Motif
To further identify potential motifs of DNA fragments recognized by CasR, a DNase I footprinting assay was performed. CasR protein was co-incubated with the DNA probe casRp, and then digested with DNase I. The results showed that the sequence AATAAGTAAGGCTTGTGTCTCACTATGA, located in the region −70 to −40 on the coding strand, was most significantly protected. In this sequence, a palindromic sequence motif was found to be separated by two partially matched inverted repeats (Figure 4A,B), suggesting that CasR recognized a palindrome sequence motif. Further EMSAs confirmed that a 20 bp motif, AGTAAGGCTTGTGTCTCACT of this sequence (AATAAGTAAGGCTTGTGTCTCACTATGA), was required for recognition by CasR. Four 28 bp DNA probe fragments, designated as casRp1–casRp4, where p1 is native DNA and p2–4 are mutants, were synthesized and subjected to EMSAs (Figure 4C). As shown in Figure 4D, CasR did not bind to the casRp4 fragments with two mutated inverted repeats; however, it bound to casRp2 and casRp3 probes with one intact inverted repeat, although the binding was slightly weaker than that to casRp1. These data indicate that half of the palindromic sequence is sufficient for CasR binding. Taken together, our findings indicate that CasR recognized a 20 bp palindrome sequence motif located in the upstream regulatory region of the casR promoter.
3.4. CasR Binds to csm6 Promoter in the Same Manner as Its Binding to Self-Promoter
In order to characterize the conserved sequence of CasR binding to the promoter of the csm6, we analyzed putative −35 (TTGAAG) and −10 (ATATGC) sequences upstream of the annotated translation start codon, based on transcription start site (Figure 5A). The results showed that the binding region of ribosomal protein was far from the translation start site. CasR-bound palindromic sequence alignment showed that there was a highly similar inverted repeat sequence in the region −331 to −301 of the csm6 promoter (Figure 5A). We speculated that CasR might regulate the expression of csm6 by directly binding to this region (−331 to −301). To test this speculation, we directly synthesized two 29 bp DNA probes, with or without IRs sequence, and performed the EMSA analysis. As shown in Figure 5B, CasR bound to csm6p1 but failed to bind to csm6p2. These results indicate that the region (−331 to −301) where CasR bound to the csm6 promoter was far from the transcription start site, and the sequence characteristics of this region are consistent with those of the CasR-binding motif.
3.5. CasR Negatively Regulates Expression of Gene csm6
To investigate the regulation of csm6 expression, we amplified a series of promoter fragments and cloned them into a promoter-less lacZ reporter plasmid and transformed these plasmids into M. smegmatis. The β-galactosidase activity analysis results showed that the strong promoter hsp60 significantly promoted the expression of lacZ, and it exhibited higher enzymatic activity than the promoter-less lacZ plasmid, indicating that the reporter system worked well (Figure 6A). Furthermore, the casRp and csm6p promoters also significantly promoted the expression of lacZ (~949.3 and 738.6 Miller units). Notably, when the open reading frame (ORF) of casR was cloned behind the promoter, the enzymatic activity of casRp-lacZ and csm6p-lacZ was significantly reduced by about 10-fold (~97.1 and 75.1 Miller units). However, when the palindromic sequence motifs of casRpm and csm6pm were mutated and used as promoters, the expression level of lacZ in the mutant strain was drastically rescued when they had the casR with the mutated promoter (~480.3 and 391.0 Miller units) (Figure 6A). These results indicate that the transcription of csm6 was suppressed by CasR.
The regulatory effect of CasR on csm6 was further examined in the M. tuberculosis H37Ra strain, and the expression of CRISPR-associated genes was quantified with qRT-PCR. When the casR gene was overexpressed via the pMV261 vector with L5 regulatory promoter (Figure 6B), the expression of casR increased by more than 12-fold in the casR-overexpressing strain relative to the pMV261 strain, indicating that the casR gene was overexpressed successfully. In contrast, csm6 expression significantly decreased by at least 2.5-fold in the casR-overexpressing strain. In addition, we detected the expression levels of other CRISPR-associated genes and found that the expression of cas1 adjacent to the downstream of csm6 was also downregulated (Figure 6B), but the expression of the distant cas2 and upstream genes of csm6 were not significantly changed, which was consistent with the EMSA results (Figure 6B).
Overall, our results suggested that CasR functioned as a transcriptional repressor, and that it negatively regulated the expressions of csm6 and cas1, rather than other CRISPR-associated genes.
4. Discussion
The CRISPR-Cas system, as an adaptive immune system in many bacteria and archaea, can defend against the invasion of foreign nucleic acids, and this system has multiple functions such as gene expression regulation, genome evolution, and DNA repair [34,35]. The expression regulation of CRISPR-Cas system genes is essential for the functional exertion of this system. However, our knowledge of the expression regulation of CRISPR-Cas system genes is still limited. In this study, we found that transcription factor CasR specifically bound to the promoter of csm6 and negatively regulated its expression.
The TetR family of transcription factors can regulate the expression of the genes related to diverse physiological functions, such as drug efflux pumps, catabolic pathway enzyme synthesis, antibiotic biosynthesis, osmotic stress, and the formation of pathogenic bacterial biofilms in bacteria [36,37,38,39,40]. In the current study, we identified and characterized a TetR family transcription factor CasR in M. tuberculosis H37Ra, and CasR was highly conserved in the strains of the M. tuberculosis complex. Our EMSA and ChIP results confirmed that CasR can bind its own promoter specifically, and it can also interact with the csm6 promoter. It is noteworthy that the binding activity of CasR to its own promoter is much stronger than to the promoter of the csm6 gene. In the EMSA experiment, the amount of CasR protein required to fully bind to its own promoter is about 1 μM, while 2 μM of the protein was needed to bind with the promoter of csm6 (Figure 3B). In addition, our mutation analysis of the CasR-binding DNA motif showed that only one shifted band was detected using casRp1 and csm6p1 as probes, indicating that a single binding motif of the casR promoter is sufficient for CasR binding; however, this may not be true for the scenario of the csm6 promoter (Figure 4D and Figure 5B). These results imply the presence of a cooperative binding effect, which has also been found for other TetR family regulators [27,36].
Our β-galactosidase analysis showed that CasR negatively regulated its own promoter expression and suppressed the expression of the CRISPR-associated gene csm6 (Figure 6A). Our qRT-PCR results confirmed that the expression level of csm6 was significantly reduced in the casR-overexpressing strain and that the expression of gene cas1 was also reduced by about two-fold (Figure 6B). The binding site of CasR within its own promoter DNA is much closer to its coding sequence, compared with that of csm6. Furthermore, sequence alignment showed that palindromic sequences of casR and csm6 promoters were highly conserved in M. tuberculosis H37Rv, M. tuberculosis H37Ra, and M. bovis BCG strains. Usually, the site recognized by RNA polymerase is near the −35 region, and our data showed that the palindromic sequence of CasR-binding csm6 promoter was located upstream of the −35 region (Figure 5A). Therefore, CasR might prevent RNA polymerase from binding to the promoter region of csm6, thus resulting in the downregulation of gene expression. Although the ChIP-seq screening revealed that the region of CasR binding to the csm6 gene was located in the coding region, we proved that its binding site was in the promoter region of csm6 by both in vitro and in vivo experiments. This indicated that there are still uncertainties in the experimental results of ChIP-seq, and the results need to be verified by further experiments. We also analyzed whether there were CasR-binding palindrome sequences in the coding sequences of the csm6 gene, and found only half a palindrome structure in the positions 994–1001. We hypothesized that CasR might also be able to weakly bind to the coding region of the csm6 gene. The binding of CasR in the csm6 coding region might act as a transcription roadblock and could also physically interfere with the transcription of cas1. The molecular mechanism by which CasR regulates the expression of CRISPR-Cas system genes can be further explored by analyzing the crystal structure of the CasR-csm6 promoter complex.
The regulation of cas gene expression is particularly important for avoiding the over-immunity of the CRISPR-Cas system after bacteria clear the invaded nucleic acids. CnpB has been reported to regulate the expression of CRISPR-Cas system genes through nanoRNA, and CnpB upregulates the expression of all the cas genes globally [22], which was inconsistent with our results that CasR accurately regulated the expression level of csm6 by binding to the promoter of the CRISPR system. In E. coli, the DNA-binding protein H-NS suppresses CRISPR-cas gene expression due to the fact that the H-NS binding sites are located near the cas operon promoters [41]. In Salmonella enterica Serovar Typhi, LRP is also a negative regulator for cas expression, with a fourfold inhibitory effect on the casA promoter [42]. Likewise, our findings indicate that CasR significantly inhibits the expression of the csm6 gene. This accurate regulation was important for bacterial response to phage- or transposon- induced horizontal gene transfer, conjugation, and transduction of plasmids carrying resistance genes. However, how CasR receives upstream signals to regulate the expression of cas genes deserves to be further investigated. We hypothesized that after phage and plasmid removal, CasR could rapidly inhibit the expression of the csm6 gene, in order to avoid the over-inhibition of bacterial growth. Based on the results of the overexpressing strain, we speculate that the mutation of casR might upregulate the gene expression of the CRISPR-Cas system. The regulable CRISPR-Cas system is necessary for the survival of bacteria, since it can save the cost of expressing a large number of cells and avoid excessive nuclease accumulation, thus enhancing bacterial immunity against foreign nucleic acid invasion.
In conclusion, this study confirmed the interaction of the TetR family CasR with its own promoter and the upstream DNA sequence of the CRISPR-Cas gene cluster identified the conserved DNA sequences recognized by CasR and preliminarily elucidated the regulatory effect of CasR on cas gene expression. Our findings provide valuable references for further studies on gene expression regulation in the CRISPR-Cas system.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/biom13020400/s1: Figure S1: SDS-PAGE of purified proteins; Figure S2: EMSA assays for the binding of Rv1255c to promoters; Figure S3: EMSA assays for the binding of Rv1473A to promoters; Figure S4: EMSA assays for the binding of CasR to promoters; Figure S5: The amino acid sequence alignment of CasR; Table S1: Bacterial strains and plasmids used in this study; Table S2: Primers used in this study.
Author Contributions
Conceptualization, M.Y.; data curation, C.-H.G.; formal analysis, W.W., X.J. and L.Z.; funding acquisition, M.Y.; investigation, W.W., X.J. and L.Z.; project administration, M.Y.; resources, M.Y.; visualization, W.W.; writing—original draft, W.W.; and writing—review and editing, W.W., Y.Y., J.Y., L.X., C.-H.G. and M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Natural Science Foundation of China (32070079 and 32100090) and the Fundamental Research Funds for the Central Universities (2020kfyXJJS119 and 2662022ZHQD001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
Much gratitude goes to linguistics Ping Liu from Huazhong Agriculture University, Wuhan, China, for her English editing and language polishing work.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Makarova, K.S.; Wolf, Y.I.; Iranzo, J.; Shmakov, S.A.; Alkhnbashi, O.S.; Brouns, S.J.J.; Charpentier, E.; Cheng, D.; Haft, D.H.; Horvath, P.; et al. Evolutionary classification of CRISPR-Cas systems: A burst of class 2 and derived variants. Nat. Rev. Microbiol. 2020, 18, 67–83. [Google Scholar] [CrossRef]
- Makarova, K.S.; Wolf, Y.I.; Koonin, E.V. Comparative genomics of defense systems in archaea and bacteria. Nucleic Acids Res. 2013, 41, 4360–4377. [Google Scholar] [CrossRef]
- Shmakov, S.; Smargon, A.; Scott, D.; Cox, D.; Pyzocha, N.; Yan, W.; Abudayyeh, O.O.; Gootenberg, J.S.; Makarova, K.S.; Wolf, Y.I.; et al. Diversity and evolution of class 2 CRISPR-Cas systems. Nat. Rev. Microbiol. 2017, 15, 169–182. [Google Scholar] [CrossRef] [Green Version]
- Weinberger, A.D.; Gilmore, M.S. A CRISPR view of cleavage. Cell 2015, 161, 964–966. [Google Scholar] [CrossRef] [Green Version]
- Makarova, K.S.; Wolf, Y.I.; Alkhnbashi, O.S.; Costa, F.; Shah, S.A.; Saunders, S.J.; Barrangou, R.; Brouns, S.J.; Charpentier, E.; Haft, D.H.; et al. An updated evolutionary classification of CRISPR-Cas systems. Nat. Rev. Microbiol. 2015, 13, 722–736. [Google Scholar] [CrossRef] [Green Version]
- Nussenzweig, P.M.; Marraffini, L.A. Molecular Mechanisms of CRISPR-Cas Immunity in Bacteria. Annu. Rev. Genet. 2020, 54, 93–120. [Google Scholar] [CrossRef]
- Strotskaya, A.; Savitskaya, E.; Metlitskaya, A.; Morozova, N.; Datsenko, K.A.; Semenova, E.; Severinov, K. The action of Escherichia coli CRISPR-Cas system on lytic bacteriophages with different lifestyles and development strategies. Nucleic Acids Res. 2017, 45, 1946–1957. [Google Scholar]
- Brouns, S.J.; Jore, M.M.; Lundgren, M.; Westra, E.R.; Slijkhuis, R.J.; Snijders, A.P.; Dickman, M.J.; Makarova, K.S.; Koonin, E.V.; van der Oost, J. Small CRISPR RNAs guide antiviral defense in prokaryotes. Science 2008, 321, 960–964. [Google Scholar] [CrossRef] [Green Version]
- Guo, T.W.; Bartesaghi, A.; Yang, H.; Falconieri, V.; Rao, P.; Merk, A.; Eng, E.T.; Raczkowski, A.M.; Fox, T.; Earl, L.A.; et al. Cryo-EM structures reveal mechanism and inhibition of DNA targeting by a CRISPR-Cas surveillance complex. Cell 2017, 171, 414–426. [Google Scholar] [CrossRef] [Green Version]
- Sternberg, S.H.; Richter, H.; Charpentier, E.; Qimron, U. Adaptation in CRISPR-Cas systems. Mol. Cell 2016, 61, 797–808. [Google Scholar] [CrossRef] [Green Version]
- Westra, E.R.; Swarts, D.C.; Staals, R.H.; Jore, M.M.; Brouns, S.J.; van der Oost, J. The CRISPRs, they are a-changin’: How prokaryotes generate adaptive immunity. Annu. Rev. Genet. 2012, 46, 311–339. [Google Scholar] [CrossRef]
- Freidlin, P.J.; Nissan, I.; Luria, A.; Goldblatt, D.; Schaffer, L.; Kaidar-Shwartz, H.; Chemtob, D.; Dveyrin, Z.; Head, S.R.; Rorman, E. Structure and variation of CRISPR and CRISPR-flanking regions in deleted-direct repeat region Mycobacterium tuberculosis complex strains. BMC Genom. 2017, 18, 168. [Google Scholar] [CrossRef] [Green Version]
- Pourcel, C.; Touchon, M.; Villeriot, N.; Vernadet, J.P.; Couvin, D.; Toffano-Nioche, C.; Vergnaud, G. CRISPRCasdb a successor of CRISPRdb containing CRISPR arrays and cas genes from complete genome sequences, and tools to download and query lists of repeats and spacers. Nucleic Acids Res. 2020, 48, D535–D544. [Google Scholar] [CrossRef]
- Tamulaitis, G.; Venclovas, C.; Siksnys, V. Type III CRISPR-Cas immunity: Major differences brushed aside. Trends Microbiol. 2017, 25, 49–61. [Google Scholar] [CrossRef]
- Supply, P.; Marceau, M.; Mangenot, S.; Roche, D.; Rouanet, C.; Khanna, V.; Majlessi, L.; Criscuolo, A.; Tap, J.; Pawlik, A.; et al. Genomic analysis of smooth tubercle bacilli provides insights into ancestry and pathoadaptation of Mycobacterium tuberculosis. Nat. Genet. 2013, 45, 172–179. [Google Scholar]
- Wei, W.; Zhang, S.; Fleming, J.; Chen, Y.; Li, Z.; Fan, S.; Liu, Y.; Wang, W.; Wang, T.; Liu, Y.; et al. Mycobacterium tuberculosis type III-A CRISPR/Cas system crRNA and its maturation have atypical features. FASEB J. 2019, 33, 1496–1509. [Google Scholar] [CrossRef]
- Gruschow, S.; Athukoralage, J.S.; Graham, S.; Hoogeboom, T.; White, M.F. Cyclic oligoadenylate signalling mediates Mycobacterium tuberculosis CRISPR defence. Nucleic Acids Res. 2019, 47, 9259–9270. [Google Scholar] [CrossRef] [Green Version]
- Jiang, W.; Samai, P.; Marraffini, L.A. Degradation of phage transcripts by CRISPR-associated RNases enables type III CRISPR-Cas immunity. Cell 2016, 164, 710–721. [Google Scholar] [CrossRef] [Green Version]
- Athukoralage, J.S.; Graham, S.; Grüschow, S.; Rouillon, C.; White, M.F. A type III CRISPR ancillary ribonuclease degrades its cyclic oligoadenylate activator. J. Mol. Biol. 2019, 431, 2894–2899. [Google Scholar] [CrossRef]
- Athukoralage, J.S.; Rouillon, C.; Graham, S.; Gruschow, S.; White, M.F. Ring nucleases deactivate type III CRISPR ribonucleases by degrading cyclic oligoadenylate. Nature 2018, 562, 277–280. [Google Scholar] [CrossRef] [Green Version]
- Richter, C.; Chang, J.T.; Fineran, P.C. Function and regulation of clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR associated (Cas) systems. Viruses 2012, 4, 2291–2311. [Google Scholar] [CrossRef]
- Zhang, Y.; Yang, J.; Bai, G. Regulation of the CRISPR-associated genes by Rv2837c (CnpB) via an orn-like activity in tuberculosis complex mycobacteria. J. Bacteriol. 2018, 200, e00743–e00817. [Google Scholar] [CrossRef] [Green Version]
- Wei, W.; Qiao, J.; Jiang, X.; Cai, L.; Hu, X.; He, J.; Chen, M.; Yang, M.; Cui, T. Dehydroquinate synthase directly binds to streptomycin and regulates susceptibility of Mycobacterium bovis to streptomycin in a non-canonical mode. Front. Microbiol. 2022, 13, 818881. [Google Scholar] [CrossRef]
- Yang, M.; Gao, C.; Cui, T.; An, J.; He, Z.G. A TetR-like regulator broadly affects the expressions of diverse genes in Mycobacterium smegmatis. Nucleic Acids Res. 2012, 40, 1009–1020. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Gao, C.H.; Hu, J.; Zhao, L.; Huang, Q.; He, Z.G. InbR, a TetR family regulator, binds with isoniazid and influences multidrug resistance in Mycobacterium bovis BCG. Sci. Rep. 2015, 5, 13969. [Google Scholar] [CrossRef] [Green Version]
- Zhu, C.; Liu, Y.; Hu, L.; Yang, M.; He, Z.G. Molecular mechanism of the synergistic activity of ethambutol and isoniazid against Mycobacterium tuberculosis. J. Biol. Chem. 2018, 293, 16741–16750. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.H.; Yang, M.; He, Z.G. Characterization of a novel ArsR-like regulator encoded by Rv2034 in Mycobacterium tuberculosis. PLoS ONE 2012, 7, e36255. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Cortes, T.; Schubert, O.T.; Rose, G.; Arnvig, K.B.; Comas, I.; Aebersold, R.; Young, D.B. Genome-wide mapping of transcriptional start sites defines an extensive leaderless transcriptome in Mycobacterium tuberculosis. Cell Rep. 2013, 5, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Jaini, S.; Lyubetskaya, A.; Gomes, A.; Peterson, M.; Tae Park, S.; Raman, S.; Schoolnik, G.; Galagan, J. Transcription factor binding site mapping using ChIP-Seq. Microbiol. Spectr. 2014, 2, 2. [Google Scholar] [CrossRef] [Green Version]
- Galagan, J.E.; Minch, K.; Peterson, M.; Lyubetskaya, A.; Azizi, E.; Sweet, L.; Gomes, A.; Rustad, T.; Dolganov, G.; Glotova, I.; et al. The Mycobacterium tuberculosis regulatory network and hypoxia. Nature 2013, 499, 178–183. [Google Scholar] [CrossRef] [Green Version]
- Gao, C.H.; Yang, M.; He, Z.G. An ArsR-like transcriptional factor recognizes a conserved sequence motif and positively regulates the expression of phoP in mycobacteria. Biochem. Biophys. Res. Commun. 2011, 411, 726–731. [Google Scholar] [CrossRef]
- Sherman, D.R.; Voskuil, M.; Schnappinger, D.; Liao, R.; Harrell, M.I.; Schoolnik, G.K. Regulation of the Mycobacterium tuberculosis hypoxic response gene encoding alpha -crystallin. Proc. Natl. Acad. Sci. USA 2001, 98, 7534–7539. [Google Scholar] [CrossRef] [Green Version]
- Mogila, I.; Kazlauskiene, M.; Valinskyte, S.; Tamulaitiene, G.; Tamulaitis, G.; Siksnys, V. Genetic dissection of the type III-A CRISPR-Cas system Csm complex reveals roles of individual subunits. Cell Rep. 2019, 26, 2753–2765.e2754. [Google Scholar] [CrossRef] [Green Version]
- Westra, E.R.; Buckling, A.; Fineran, P.C. CRISPR-Cas systems: Beyond adaptive immunity. Nat. Rev. Microbiol. 2014, 12, 317–326. [Google Scholar] [CrossRef]
- Ramos, J.L.; Martínez-Bueno, M.; Molina-Henares, A.J.; Terán, W.; Watanabe, K.; Zhang, X.; Gallegos, M.T.; Brennan, R.; Tobes, R. The TetR family of transcriptional repressors. Microbiol. Mol. Biol. Rev. 2005, 69, 326–356. [Google Scholar] [CrossRef] [Green Version]
- Balhana, R.J.; Singla, A.; Sikder, M.H.; Withers, M.; Kendall, S.L. Global analyses of TetR family transcriptional regulators in mycobacteria indicates conservation across species and diversity in regulated functions. BMC Genom. 2015, 16, 479. [Google Scholar] [CrossRef] [Green Version]
- Miotto, P.; Sorrentino, R.; De Giorgi, S.; Provvedi, R.; Cirillo, D.M.; Manganelli, R. Transcriptional regulation and drug resistance in Mycobacterium tuberculosis. Front. Cell. Infect. Microbiol. 2022, 12, 990312. [Google Scholar] [CrossRef]
- Wang, K.; Sybers, D.; Maklad, H.R.; Lemmens, L.; Lewyllie, C.; Zhou, X.; Schult, F.; Bräsen, C.; Siebers, B.; Valegård, K.; et al. A TetR-family transcription factor regulates fatty acid metabolism in the archaeal model organism Sulfolobus acidocaldarius. Nat. Commun. 2019, 10, 1542. [Google Scholar] [CrossRef] [Green Version]
- Carette, X.; Blondiaux, N.; Willery, E.; Hoos, S.; Lecat-Guillet, N.; Lens, Z.; Wohlkönig, A.; Wintjens, R.; Soror, S.H.; Frénois, F.; et al. Structural activation of the transcriptional repressor EthR from Mycobacterium tuberculosis by single amino acid change mimicking natural and synthetic ligands. Nucleic Acids Res. 2012, 40, 3018–3030. [Google Scholar] [CrossRef] [Green Version]
- Pul, U.; Wurm, R.; Arslan, Z.; Geissen, R.; Hofmann, N.; Wagner, R. Identification and characterization of E. coli CRISPR-cas promoters and their silencing by H-NS. Mol. Microbiol. 2010, 75, 1495–1512. [Google Scholar] [CrossRef]
- Medina-Aparicio, L.; Rebollar-Flores, J.E.; Gallego-Hernández, A.L.; Vázquez, A.; Olvera, L.; Gutiérrez-Ríos, R.M.; Calva, E.; Hernández-Lucas, I. The CRISPR/Cas immune system is an operon regulated by LeuO, H-NS, and leucine-responsive regulatory protein in Salmonella enterica serovar Typhi. J. Bacteriol. 2011, 193, 2396–2407. [Google Scholar] [CrossRef] [Green Version]
Figure 1. Type III-A CRISPR system of M. tuberculosis. (A) Genetic organization of the CRISPR-Cas system in M. tuberculosis. In M. tuberculosis, the CRISPR-Cas system is a type III-A system, and the expression pattern may differ from that of other cas genes. An independent transcription start site (TSS) was identified upstream of the gene csm6 (red arrow). The TSS of other genes are indicated with black arrows in the schematic. (B) Predicted transcription factors interacting with Csm6. The blocks in purple refer to the predicted transcription factors, and the locations indicate the maximum coverage of possible binding sites. Fold change is the expression of a target gene (csm6) when the corresponding transcription factor is induced, as noted in the TB database. Binding types are either genic or intergenic, based on the location of maximum coverage for the binding site relative to predicted genes.
Figure 1. Type III-A CRISPR system of M. tuberculosis. (A) Genetic organization of the CRISPR-Cas system in M. tuberculosis. In M. tuberculosis, the CRISPR-Cas system is a type III-A system, and the expression pattern may differ from that of other cas genes. An independent transcription start site (TSS) was identified upstream of the gene csm6 (red arrow). The TSS of other genes are indicated with black arrows in the schematic. (B) Predicted transcription factors interacting with Csm6. The blocks in purple refer to the predicted transcription factors, and the locations indicate the maximum coverage of possible binding sites. Fold change is the expression of a target gene (csm6) when the corresponding transcription factor is induced, as noted in the TB database. Binding types are either genic or intergenic, based on the location of maximum coverage for the binding site relative to predicted genes.

Figure 2. EMSA of binding of transcription factors (A) Rv0324, (B) Rv1255c, and (C) Rv1473A to their own promoters and CRISPR system promoters. The 100 ng target gene csm6 and 100 ng cas6 promoters were co-incubated with different concentrations (0–2 μM) of transcription factors. The DNA-protein complex is indicated with square brackets.
Figure 2. EMSA of binding of transcription factors (A) Rv0324, (B) Rv1255c, and (C) Rv1473A to their own promoters and CRISPR system promoters. The 100 ng target gene csm6 and 100 ng cas6 promoters were co-incubated with different concentrations (0–2 μM) of transcription factors. The DNA-protein complex is indicated with square brackets.

Figure 3. Specific interaction of CasR with the promoter of the csm6 in CRISPR-Cas system of M. tuberculosis. (A) Schematic diagram of the promoter fragment used in this experiment. (B) EMSA of CasR binding to its own promoter and CRISPR system promoters. The 100 ng casR promoter (casRp, Lanes 1–4) and 100 ng target gene csm6 promoter (csm6p, Lanes 5–8) were incubated with different concentrations (0–2 μM) of CasR protein. The non-specific promoter cas6p (Lanes 9–12) served as a negative control. The DNA–protein complex is indicated with square brackets. (C) ChIP assay in the wild-type M. tuberculosis H37Ra strain. In ChIP, P indicates pre-immune serum, and I denotes antiserum against CasR. The mycobacterial promoter cas6p was used as a negative control. (D) CasR amino acid sequence with 100% identity in M. tuberculosis H37Rv, M. tuberculosis H37Ra, and M. bovis BCG. (E) Domain characteristics of CasR. CasR contained an HTH TetR_type superfamily domain and a C-terminal domain with unknown function.
Figure 3. Specific interaction of CasR with the promoter of the csm6 in CRISPR-Cas system of M. tuberculosis. (A) Schematic diagram of the promoter fragment used in this experiment. (B) EMSA of CasR binding to its own promoter and CRISPR system promoters. The 100 ng casR promoter (casRp, Lanes 1–4) and 100 ng target gene csm6 promoter (csm6p, Lanes 5–8) were incubated with different concentrations (0–2 μM) of CasR protein. The non-specific promoter cas6p (Lanes 9–12) served as a negative control. The DNA–protein complex is indicated with square brackets. (C) ChIP assay in the wild-type M. tuberculosis H37Ra strain. In ChIP, P indicates pre-immune serum, and I denotes antiserum against CasR. The mycobacterial promoter cas6p was used as a negative control. (D) CasR amino acid sequence with 100% identity in M. tuberculosis H37Rv, M. tuberculosis H37Ra, and M. bovis BCG. (E) Domain characteristics of CasR. CasR contained an HTH TetR_type superfamily domain and a C-terminal domain with unknown function.

Figure 4. Profile of motif sequence recognized by CasR. (A) DNase I footprinting assay. The protective effect of CasR on the promoter DNA against DNase I digestion increased with increasing concentrations of CasR (0 μM, 1.2 μM). The sequences in the protected regions on the coding strand are underlined in the figures. (B) Sequence and structural characteristics of the promoter region protected by CasR. The regions protected by CasR are indicated with underlines, and the box indicates the 20 bp sequences containing two inverted repeats (IRs) with a 4 bp interval. The translation start codon of CasR is indicated in bold. (C) EMSA of DNA sequences with different mutations. Three mutant DNA probes contain left part mutation, right part mutation, and complete mutation of palindromic structure. (D) EMSA of four mutant DNA probes, with or without the IR sequence. Each DNA probe was co-incubated with 0–2 μM CasR protein.
Figure 4. Profile of motif sequence recognized by CasR. (A) DNase I footprinting assay. The protective effect of CasR on the promoter DNA against DNase I digestion increased with increasing concentrations of CasR (0 μM, 1.2 μM). The sequences in the protected regions on the coding strand are underlined in the figures. (B) Sequence and structural characteristics of the promoter region protected by CasR. The regions protected by CasR are indicated with underlines, and the box indicates the 20 bp sequences containing two inverted repeats (IRs) with a 4 bp interval. The translation start codon of CasR is indicated in bold. (C) EMSA of DNA sequences with different mutations. Three mutant DNA probes contain left part mutation, right part mutation, and complete mutation of palindromic structure. (D) EMSA of four mutant DNA probes, with or without the IR sequence. Each DNA probe was co-incubated with 0–2 μM CasR protein.

Figure 5. Motif of CasR-binding csm6 promoter. (A) Sequence and structural characterization of CasR-protected csm6 promoter region. The CasR-protected regions from −331 to −306 are indicated with underlines. Putative −10 and −35 sequences are marked. The approximate transcription start site at position −132 is marked with TSS. (B) EMSA of DNA-binding activity of CasR on the DNA probes, with (csm6p1, Lanes 1–4) or without IR sequence (csm6p2, Lanes 5–8). Either DNA probe was co-incubated with 0–2 μM CasR protein.
Figure 5. Motif of CasR-binding csm6 promoter. (A) Sequence and structural characterization of CasR-protected csm6 promoter region. The CasR-protected regions from −331 to −306 are indicated with underlines. Putative −10 and −35 sequences are marked. The approximate transcription start site at position −132 is marked with TSS. (B) EMSA of DNA-binding activity of CasR on the DNA probes, with (csm6p1, Lanes 1–4) or without IR sequence (csm6p2, Lanes 5–8). Either DNA probe was co-incubated with 0–2 μM CasR protein.

Figure 6. CasR negatively regulates the expression of csm6. (A) β-galactosidase activity assay. Schematic diagram of plasmids containing promoter-lacZ and promoter-CasR ORF-lacZ. All the plasmids were transformed into the M. smegmatis. The activity of β-galactosidase is expressed as Miller units. Data are expressed as the mean ± standard deviations of three independent replicates. (B) Relative expression levels of CRISPR-associated genes via qRT-PCR in the casR-overexpressing strain and pMV261 control strain. The relative expression levels of the genes were normalized using the sigA gene as internal control. Relative expression levels of genes were calculated using the 2−ΔΔCt method. The differences in the relative expression levels between groups were analyzed via unpaired two-tailed Student’s t-test, using GraphPad Prism 8. Asterisks represent significant differences (***, p < 0.001) between two groups.
Figure 6. CasR negatively regulates the expression of csm6. (A) β-galactosidase activity assay. Schematic diagram of plasmids containing promoter-lacZ and promoter-CasR ORF-lacZ. All the plasmids were transformed into the M. smegmatis. The activity of β-galactosidase is expressed as Miller units. Data are expressed as the mean ± standard deviations of three independent replicates. (B) Relative expression levels of CRISPR-associated genes via qRT-PCR in the casR-overexpressing strain and pMV261 control strain. The relative expression levels of the genes were normalized using the sigA gene as internal control. Relative expression levels of genes were calculated using the 2−ΔΔCt method. The differences in the relative expression levels between groups were analyzed via unpaired two-tailed Student’s t-test, using GraphPad Prism 8. Asterisks represent significant differences (***, p < 0.001) between two groups.

|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content.
|
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Wei, W.; Jiang, X.; Zhang, L.; Yan, Y.; Yan, J.; Xu, L.; Gao, C.-H.; Yang, M. Regulation of CRISPR-Associated Genes by Rv1776c (CasR) in Mycobacterium tuberculosis. Biomolecules 2023, 13, 400. https://doi.org/10.3390/biom13020400
AMA Style
Wei W, Jiang X, Zhang L, Yan Y, Yan J, Xu L, Gao C-H, Yang M. Regulation of CRISPR-Associated Genes by Rv1776c (CasR) in Mycobacterium tuberculosis. Biomolecules. 2023; 13(2):400. https://doi.org/10.3390/biom13020400
Chicago/Turabian StyleWei, Wenping, Xiaofang Jiang, Li Zhang, Yunjun Yan, Jinyong Yan, Li Xu, Chun-Hui Gao, and Min Yang. 2023. "Regulation of CRISPR-Associated Genes by Rv1776c (CasR) in Mycobacterium tuberculosis" Biomolecules 13, no. 2: 400. https://doi.org/10.3390/biom13020400
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.