
Abstract
With the unexpected emergence of the novel 2019 Wuhan coronavirus, the world was faced with a sudden uproar that quickly shifted into a serious life-threatening pandemic. Affecting the lives of the global population and leaving drastic damage in various sections and systems, several measures have been constantly taken to tackle down this crisis. For instance, numerous vaccines have been developed in the past two years, some of which have been granted emergency use, thus providing sufficient immunity to the vaccinated individuals. However, the appearance of newly emerged SARS-CoV-2 variants with accelerated transmission and fatality has led the world towards another pandemic. Having undergone various mutations in genomic and/or amino acid profiles, some of the emerged variants of concern (VOCs) including Alpha, Beta, Gamma, and Delta have displayed immune evasion and pathogenicity even in the vaccinated population, hence raising concerns regarding the efficacy of current vaccines against new VOCs of COVID-19. Therefore, genomic investigations of SARS-CoV-2 mutations are expected to provide valuable insight into the evolution of SARS-CoV-2, while also determining the impact of different mutations on infection severity. This study was constructed with the aim of shining light on recent advances regarding mutations in major COVID-19 VOCs, as well as vaccination efficacy against those VOCs.
Similar content being viewed by others
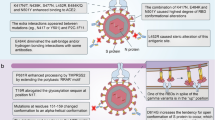


Introduction
Ever since late 2019, the rapid emergence of a novel coronavirus named SARS-CoV-2 or COVID-19 has resulted in a life-threatening pandemic situation. Even though originally found in Wuhan province of China, COVID-19 has managed to spread globally at a fascinating rate thus resulting in a worldwide outbreak [1]. Owing to its incredibly fast spread and significant mortality rates, the new SARS-CoV-2 has continuously endangered the lives of many and has caused millions of deaths all around the globe [2, 3].
This non-segmented, positive-sense, enveloped virus possesses single-stranded RNA possesses an ever-evolving nature that has resulted in the emergence of numerous viral variants with alterations in genomic or amino acid profiles [4]. For instance, some of the mutations that take place in the Spike glycoprotein of SARS-CoV-2 enable the virus to escape the host’s immune system or increase the binding affinity of Spike’s receptor-binding domain (RBD) to its host Angiotensin-converting enzyme II (ACE2) receptors, hence alter some of the viral characteristics, such as increase viral pathogenicity and subsequent infection severity [5]. Moreover, mutations in the viral genome and amino acid profiles are also a subject of major research interest, due to their evident impact on vaccination efficacies [6,7,8,9].
While global vaccination is considered to be the golden approach for controlling the pandemic, it is crucial to note that formulation, development, and industrial production of vaccines is a task with great difficulty [10]. With the employment of various traditional and novel platforms for vaccine development, there is now a tremendous load of information regarding this highly complex process as well as its effectiveness against SARS-CoV-2. On the other hand, the emergence of new COVID-19 variants has recently raised a lot of concerns regarding the current vaccination efficacy against new variants [11, 12]. Even though data in this respective area is now growing at an ever-increasing speed, current literature is still faced with a lack of sufficient cohesion in available data. Therefore, this review was conducted with aim of providing well-organized information regarding SARS-CoV-2 genome analysis and current vaccine platforms with a special focus on the novel Delta (B.1.617.2) variant. It is in this work’s best hopes to provide deep insight on current advances in this regard, as well as further elucidate the path to conducting new experiments and overcoming this ongoing battle of humans against coronaviruses.
Discussion
An overview on SARS-CoV-2: What we currently know
Clinical presentations and diagnostic tools
Individuals infected with COVID-19 might experience various symptoms. Fever, dry coughs, and shortness of breath are by far the most common symptoms observed in 83%, 82%, and 31% of patients, respectively [13]. Additionally, about 2–10% of patients were reported to have gastrointestinal symptoms including diarrhea, vomiting, and abdominal aches [13, 14]. Patients that experience severe symptoms are often admitted to ICU and are very likely to display symptoms of anorexia, diarrhea, dyspnea, dizziness, and muscular aches in the abdominal region [13]. In these cases, pneumonia, acute respiratory distress syndrome, multiple organ failure, and other serious conditions might occur, which are the most common causes of death due to coronavirus [15, 16].
Chest CT scans of patients with pneumonia and acute respiratory distress syndrome display ground-glass opacities that grow larger with time bizarre-paving patterns, lung consolidation, existence of nodular/cord-like shadows, and interlobular septal thickening, vascular thickening, and air broncho-gram signs inside the lesion in some cases [17]. Lesions are then absorbed after two weeks of opacity growth and left extensive opacities and subpleural parenchymal bands gradually as patients are starting to recover [18]. It is also important to mention that in the blood tests of patients with severe symptoms, leukopenia and high levels of ferritin have been observed as well as increased liver functions [19]. Of note, various investigations have aimed to explore potential factors that impact the severity of COVID-19 infection. Lymphopenia, described as reduced counts of lymphocytes in the blood, has been associated with contradictory results in the context of COVID-19 infection. However, a recent systematic review has suggested that COVID-19 infection is capable of decreasing the number of lymphocytes and impairing their normal functions, as individuals with severe COVID-19 infection had dramatically reduced lymphocyte counts in comparison with non-severe cases. Therefore, lymphocyte-targeting approaches may be of promising therapeutic value for increasing lymphocyte numbers thus improving the state of severely infected patients [20]. Furthermore, it is well-established that underlying co-morbidities such as cardiovascular diseases, diabetes, and metabolic disorders often play detrimental roles in the worsening of COVID-19 infection [21]. Metabolic dysfunctions affect many people worldwide and are considered to be a baseline of inflammation in suffering individuals [22]. Cytokine upregulations, insulin resistance, and impaired endocrine/paracrine signaling are some of the important downsides of such disorders, which may predispose patients with severe forms of COVID-19 infection [23]. On the other hand, insulin resistance has been linked with increased oxidative stress which can, in turn, expose β cells to consequent inflammation and functional impairment, hence leading to possible metabolic issues in long terms after COVID-19 infection [24, 25]. Oxidative stress, resulting from overproduction of reactive oxygen species, contributes to the progressions of a broad range of disorders such as gastrointestinal disorders [26], metabolic syndrome, and even deteriorates the severity of COVID-19 infection and is associated with cytokine storm amplification and cell hypoxia [27]. Strikingly, new studies suggest that COVID-19 may subsequently lead to the occurrence of new-onset diabetes [28] and pancreatitis [29].
RT-PCR assay is the most common method for diagnosis of COVID-19, but false-negative results are also possible in some samples [30], therefore in patients with suspicious clinical symptoms, repeated sampling is highly advised [31]. Sampling procedures are recommended to be carried out from the lower respiratory tract such as saliva and endotracheal aspirates, however, aerosols are often produced during this process and strict airborne precaution measures are highly required [32]. Moreover, bronchoalveolar lavage-based diagnosis has displayed a high yield for obtaining reliable results, but it is generally prohibited in order to minimize potential exposure risks for the healthcare team [33]. Chest CT scan is another reliable technique that was recently proved to be more efficient than RT-PCR assays [34] as it is shown to have 98% sensitivity which is significantly higher than the 71% sensitivity of the RT-PCR method [35].
Primary modeling results have displayed a log-normal distribution, which has also been observed in other acute respiratory diseases caused by viruses [36]. The average incubation period is 5.1 days, and 97.5% of patients have been reported to experience symptoms within 11.5 days post the initial infection. In an analysis based on 88 patients in Wuhan, the incubation period was indicated to be 6.4 days and changing in a range of 2.1 to 11.2 days [37, 38]. Another analysis of 158 people in Wuhan indicated a mean incubation period of 5.0 days ranging from 2 to 14 days [37]. It was also stated that 101 out of 10,000 infected patients develop symptoms after 14 days of quarantine [39], while a few cases displayed symptoms 24 days after initial infection, which is considered to be the longest incubation period yet observed. Moreover, a 14-day quarantine and monitoring are highly suggested for people who have had prior contact with potentially infected people in order to reduce the risk for further spread of this virus [37]. Interestingly, a recent study indicated that the mean duration of infection with mild or moderate COVID-19 is about 24.42 ± 1.67 days in the first episode, and reduced to 15.38 ± 5.57 days for the second episode (for mild or moderate reinfection). However, the severe form of infection lasted for an average of 21.80 ± 3.79 days and 19.20 ± 2.98 days for the first and second episode of infection, respectively [40].
Taxonomy of SARS-CoV-2 and variants of concern (VOCs)
SARS-CoV-2 is a member of the Orthocoronaviridae subfamily of the Coronaviridae family, belonging to the Nidovirales order. The large Coronoviridae family consists of four Alpha, Beta, Delta, and Gamma coronaviruses genera, among which COVID-19 falls into the Betacoronavirus genera [41]. The Betacoronivirus genus is comprised of four Embecovirus, Nobevirus, Merbecovirus, Sarbecovirus sub-genera, among which SARS-CoV-2, SARS, and Bat-SL-CoV reside in the latter sub-genera [42]. Due to the ever-evolving nature of coronaviruses and continuous emergence of new variants, the taxonomic classification of these viral species has undergone several re-examinations [41], and the final adjustment of this taxonomic classification is presented by the International Committee on Taxonomy of Viruses Executive [43].
The first variant of concern (VOC), known as the B.1.1.7 variant (Alpha VOC), was reported to possess 50–80% higher transmissibility due to the occurrence of several mutations in the RBD region of the spike glycoprotein [2, 44]. Being initially found in England, the B.1.1.7 variant managed to pave its way out to spread all around Europe in a short span of time [45]. Other VOCs were then detected in different regions, some of which were associated with increased transmissibility and mortality rates. As demonstrated in Table 1, B.1.351 VOC from South Africa [49], P.1 from Brazil [51], and B.1.617.2 variant from India (known as the novel Delta coronavirus) are some of the most fatal SARS-CoV-2 variants that have resulted in recurrent pandemic waves [51, 56] due to their accelerated contagion and drastic mortality [57].
With the constant evolution of this viral species, the emergence of novel VOCs is now a well-expected issue that has rapidly raised global concerns. As a result, genomic investigation of SARS-CoV-2 variants has recently attracted a lot of attention, as it may help to improve our understanding of SARS-CoV-2 viral origins and evolution pathways, help to detect highly mutable genomic regions, display valuable information regarding the correlation between different mutations and consequent pathogenicity and transmissibility, and also assist us in deepening our insight on the efficiency of current therapeutic approaches and vaccine development platforms against newly-emerging VOCs. For example, whole-genome analysis of SARS-CoV-2 strains has previously exhibited ~88%, ~87%, ~79%, and ~ 50% genomic similarity between the primary 2019 SARS-CoV-2 and bat-SL-CoVZC45, bat-SL-CoVZXC21, SARS-CoV, MERS-CoV, all in a respective order [58, 59]. However, the highest genomic similarity of the first Covid-19 sequence was detected in the bat-CoV (RaTG13) strain which further elucidates the possible origins of this virus [60]. Other investigations have also been conducted with the aim of studying genetic diversity between SARS-CoV-2 strains collected from multiple regions, some of which have published their obtained sequences in the GISAID database (https://www.gisaid.org/). Some of these studies specify the abundance and types of mutations (synonymous- nonsynonymous- deletion- insertion) in different genomic regions of the viral isolates [61], while some others mainly discuss high-frequency mutations across the sequenced COVID-19 strains [62] as these mutations can play a major role in viral transmissibility and pathogenicity [63].
Evolution of SARS-CoV-2: A genomic and amino acid perspective
Genomic organization of SARS-CoV-2
The genome of SARS-CoV-2 has been sequenced with metagenomic sequencing techniques and a total of 9860 amino acids were reported to be encoded by an entire genome size of 29,881 bp (NCBI genome databank, NC_045512.2) [64]. As presented in Table 2, genome compartments of SARS-CoV-2 fall into four main regions of 5′-UTR (untranslated region), 3′-UTR, non-structural regions, and structural regions [65]. To our knowledge, the initial 20 kb of the SARS-CoV-2 genome is composed of 5′-UTR and two open reading frames (ORF1a and ORF1ab). ORF1a and ORF1ab are responsible for encoding 16 non-structural proteins (nsp1–16), the first eleven of which are also called polyproteins 1a (pp1a), and the rest are called pp1ab [68]. These NSPs are in charge of a wide range of viral functions. For instance, nsp1 is known to shut off the host’s innate immunity and bind to the 40 S ribosomal complex of the host cell [69]. Nsp2 is considered to induce the production of type1 interferon (IFN-1) [70], while nsp3 and nsp5 are in charge of coding two vital proteases named papain-like protease (PLpro) and 3–chymotrypsin-like main protease(3CLpro), in a respective order [71].
Moreover, the structural genome consists of 13–15 ORFs (ORF3a, ORF3d, ORF6, ORF7a, ORF7b, ORF8, ORF9b, ORF14, and ORF10) as well as the S, E, M, and N regions [72, 73]. This region is mainly responsible for coding four crucial structural proteins of SARS-CoV-2, referred to as the Spike protein (S), Membrane (M), Envelope (E), and nucleocaspid (N) [74, 75]. Given the vital roles of ORFs and S, E, M, and N regions of SARS-CoV-2 and their consequent impact on viral pathogenicity, these genomic compartments are targets of significant value for vaccine development and drug design platforms [65].
The spike glycoprotein is a transmembrane protein that is capable of forming protruding homotrimers from the viral surface and is responsible for mediating virus invasion into host cells [76]. The S glycoprotein consists of S1 and S2 subunits that attribute to host cell receptor binding, and cellular and viral membrane fusion in a respective manner [77]. For the means of entering host cells, different coronaviruses use different domains of the S1 subunit named SA and SB domains. The SB domain is employed by SARS-CoV-2 and other SARS-related coronaviruses to interact with angiotensin-converting enzyme and subsequently enter host cells [78]. Upon the binding of S protein and host cell receptor, virus entry into host cells is further facilitated through the activation of the S protein with the help of a host cell membrane enzyme called type 2 TM serine protease (TMPRSS2). Sequential to virus entry and the release of viral RNA into host cells, genome replication and transcription processes take place right before viral structural proteins are synthesized [4].
Due to the crucial importance of these proteins especially the S glycoprotein, various therapeutic strategies have been constructed upon targeting them. For this regard, 3CLpro inhibitor (3CLpro-1), ACE2-based peptides, and vinylsulfone protease inhibitor have been suggested and used efficiently for fighting the new coronavirus [79].
SARS-CoV-2 mutations
Main genetic mutations
The rate for mutation in viral species is a multifactorial issue in which several factors including genome type (RNA or DNA) and length play an important role [75]. Generally, DNA viruses are considered to experience fewer mutations due to their shorter genome, however, most DNA-containing viruses with larger genomic contents often benefit from the existence of a DNA repairing protein that increases their chance of possessing more favorable viral mutations. On the other hand, RNA viruses often have smaller genomic contents and thus are mostly associated with a more significant mutation rate. Another factor that is positively correlated with higher mutation rates in RNA-containing viruses is an enzyme called RNA-dependent RNA polymerase (RdRp). This enzyme, however, does not possess proofreading abilities due to the absence of a 3′ exonuclease domain and is, therefore, unable to repair non-favorable genomic mutations. In contrast, Coronaviruses possess significant proofreading activities that are not dependent on their RdRp enzymes. Moreover, RdRp of Coronaviruses consists of a 3′ exonuclease domain that results in the occurrence of fewer mutations as well as a larger genome size (about 26 kb) contrary to most other RNA-containing viruses [80].
Besides virus-dependent factors that contribute to the alterations in viral mutation rates, there are various host-dependent factors including host-encoded deaminases, apolipoprotein B mRNA editing catalytic polypeptide-like enzymes (APOBEC), and Urcacil DNA glycosylases (UNG) that play a critical role in viral mutation rates [81, 82].
COVID-19 virus has been predicted to undergo various mutations at a rate of 8.90e−04 subs/site/year which is close to the mutation rate predicted for other viruses, meaning that each genome sequence in the viral genome is subject to ~26 mutations each year [83]. The reference sequence of COVID-19 has been represented by the scrutiny of genetic diversity between different strains. Several studies have investigated various SARS-CoV-2 strains and classified them based on high-frequency mutations and informative nucleotide positions in different regions of the genome [84, 85]. In a study by Tang et al. [86], strains were categorized into S and L sections based on two nucleotides of 28,144 and 8782. Zhao et al. [85], used 17 nucleotides as a basis to separate strains into 19 groups with different geographical distributions. In contradiction to the geographical distribution of the MERS virus in which a special group exists in a specific region, the geographical distribution of SARS-CoV-2 different clades revealed that different clades exist in different regions but their abundancies differ from region to region which is due to the founder effects [87]. Synonymous mutations affect translation and transcription, and nonsynonymous mutations affect the structure and functions of the related proteins. Therefore, more investigations are required to decode the exact influences of different mutations in different strains. The occurrence of mutations, especially nonsynonymous mutations with high frequency in coding regions such as the S protein, should be taken into consideration when developing new therapeutic strategies or vaccine designs [88, 89]. Several studies have investigated the functional effects of some types of mutations, for instance, mutation in the 28144th nucleotide of ORF8 results in the replacement of leucine amino acid with serine which effects the structural conformation of the protein [90]. Deletion of the 302nd nucleotide in ORF8 has resulted in an increase of N gene expression [91].
For this regard, major amino acid substitutions that have been previously detected in amino acid sequences of COVID-19’s spike protein and confirmed by FUBAR (Fast Unconstrained Bayesian Approximation) are briefly discussed in the following section [92].
Main amino acid mutations and substitutions
Spike RBD mutations
Among the different regions of SARS-CoV-2 spike protein, RBD (receptor binding domain) and NTD (N-terminal domain) are two of the most important immunologically provoking regions [93,94,95]. Therefore, any mutations in these areas that enhance viral escaping from the immune system, are highly likely to reduce antibody-mediated neutralization. For instance, E484 residue of the RBD region is known to be a critical residue that can undergo amino acid substitutions with K, Q, or P and thus decrease immune system neutralization dramatically [96]. Another escape route in times of exposure with C121 and C144 mABs is the powerful E484K mutation that is capable of diminishing neutralization effects of two REGN10989 and REGN10934 mABs [88, 97, 98]. There is also other evidence that further emphasizes the importance of the E484 mutation, as all four E484A, E484D, E484G and E484 mutations of this residue exhibited great potential in reducing immune responses of all four sera in that study. However, a few mutations such as K444E, G446V, L452R, and F490S managed to escape three of the sera in that study. The same study also detected mutations at position 477 of the S glycoprotein (S477G, S477N, and S477R) that were capable of escaping mABs, and the S477G mutation was reported to be resistant to two of the four investigated sera.
A controversial mutation for which there is still not a clear result available is the N439K mutation. A recent study by Greaney et al., reported that this mutation is not associated with any remarkable reductions in polyclonal antibody-mediated neutralization in plasma [96]. However, another study indicated that this mutation is highly likely to result in the formation of a new salt bridge between RBM-ACE2 and thus promote viral binding to ACE2, and consequently disrupt neutralization by mABs and plasma, especially in a plasma with previously lower neutralization potential [99]. Such controversy in obtained results can be attributed to variations in experimental designs, detection techniques sensitivity, and different evasion routes than what was previously predicted to be only related to reducing the recognition capacities of antibodies. K444R, K444N, K444Q, V445E, and E484K are also several other immune system evasion mutations that have occurred in RBD [88, 97].
Spike NTD mutations
N3 loops (residues 140–156) and N5 loop (246–260 residues) are two of the most critical regions of Spike’s NTD that are located at a conformational epitope and undergo mutations in order to increase viral immune escape rates [100]. There is strong evidence suggesting that deletion mutations play a continuous and remarkable role in altering NTD antigenic properties as well as contributing to the rapid transformation of COVID-19 [88, 94, 101]. RDR1 (Δ69–70), RDR2 (Δ141–144 and Δ146), RDR3 (Δ210), and RDR4 (Δ243–244) are four recurrently deleted regions (RDRs) of NTD, among which RDR1, RDR2, and RDR4 are situated in N2, N3, and N5 loops, while RDR3 is located in an area between N4 and N5 loops [101]. RDR2 deletions were shown to be associated with the emergence of Δ140 when incubated convalescent plasma, and causing a fourfold reduction in neutralization titer. Moreover, this Δ140 mutant variant resulted in the E484K mutation, which led to a further decrease in plasma neutralization titer [88]. This Δ140 + E484K mutant then caused a new insertion mutation throughout which 11 residues were added between Y248 and L249 compartments within the NTD N5 loop, followed by which, plasma neutralization was completely inhibited [88].
Furthermore, mutations at 15th and 136th cysteine residues of the signal peptide region within NTD, such as C136Y and S12P, are capable of altering the neutralization capacities of a few different mABs through the disruption of disulfide bond formation and removal of the antibody-targeted supersite [94]. ΔF140, N148S, K150R, K150E, K150T, K150Q, and S151P are also some of the other evasion mutations that have taken place in NTD [88, 97].
According to the GISAID database, mutations at 18th, 614th, and 222nd amino acid residues were reported to be the most common mutations compared to the original Wuhan-Hu-1 reference sequence (MN908947) [5]. Sequential to that, A222V is considered to be the second most common amino acid substitution in spike glycoprotein and has been found in the B.1.177 lineage which was originated from Spain [102]. Last but not least, the L18F amino acid substitution that is capable of evading from various NTD-binding mABs [94], has been reported to have occurred about 21 times globally [103].
The ever-evolving nature of the new SARS-CoV-2 has been associated with the occurrence of new mutations and variants of COVID-19, of which the novel delta (B.1.617.2) variant is a recent example. The severity of pathogenicity and transmission rates of SARS-CoV-2 are in close correlation with the number and types of genomic and amino acid substitutions that occur in the virus. Such substitutions are also capable of determining the cross-protection of previous infections and/or vaccination efficacies against new viral variants [5, 8]. This issue has recently raised a lot of doubts regarding the efficacy and viral coverage of current vaccines. On the other hand, the lack of sufficient data on potential mutations that might alter viral pathogenicity and transmissibility is another issue of major concern. Recently, a new variant of concern named Omicron (B.1.1.529) was detected in South Africa which is now rapidly spreading throughout the globe. Having up to 59 mutations throughout its genome, this variant has shifted worldwide attention due to its enhanced immune system-escaping abilities, higher reinfection risks, and increased transmissibility. Omicron VOC contains 15 mutations located in the RBD including K417, E484, and N501 which are also present in Beta and Gamma VOCs and reduce vaccine-induced neutralization [104]. A recent study has revealed that Omicron possesses higher infectivity and successfully evades current vaccination regimens, however, booster doses of current mRNA vaccines have exhibited promising results thus suggesting their potential effectiveness in overcoming immune system evasion of Omicron VOC [105, 106].
Vaccination platforms: Where we stand and what to expect
COVID-19 has undoubtedly disrupted the lives of many people all around the world. Ever since the beginning of the pandemic, more than half of the earth’s population has been going by extreme safety measures and limitations from social distancing in order to prevent further spread and infection of the virus. Under current circumstances, vaccines are of critical importance in order to provide safety and help global citizens to go back to their normal lives. To this day, various drug discovery and drug repurposing strategies [107, 108] and vaccine development approaches have been employed to enhance the public’s immunity and reduce the global burden resulting from COVID-19 [109]. Moreover, experiences from previous efforts against other coronaviruses such as MERS and SARS have substantially helped in speeding up the process of COVID-19 vaccine development. There are currently a few FDA-approved vaccines that are being extensively used to immune people in a lot of countries. While obtaining significant results regarding the efficacies of these vaccines, there is a lot of doubt regarding their efficacy against new mutations of SARS-CoV-2, especially the Delta (B.617.2) and Omicron (B.1.1.529) variants that have recently raised the question of whether the world is moving toward another pandemic [110]. The answer to this question depends on a set of different factors and falls in close correlation with whether new genomic or amino acid mutations of SARS-CoV-2 will provide it with new escape routes from vaccine-induced immunity. Answering this question requires deeper investigations and more thorough analyses. However, a comprehensive classification of current literature in regard to vaccine development and efficacy against SARS-CoV-2 is considered to be of significant importance in order to provide scientists with an inclusive insight regarding where we now stand with current vaccines, and what we might expect in the near future.
Main vaccine development platforms
Inactivated viruses
Inactivated virus-based vaccines are quite easy to obtain and can be produced on a large scale with a rather fast speed. In regard to SARS and MERS viral species, inactivated virus-based platforms have previously shown remarkable immunization, and have also been capable of triggering both humoral and cellular immunity in animal models. However, it is not clear if this stimulation of the immune system response has the ability to induce complete immunity from the disease or not. This group of vaccines might also cause unwanted immune and inflammatory responses.
Development of Sinopharm’s WBIP-CorV vaccine (Vero cells) as the first inactivated virus-based vaccine for COVID-19, was followed by the development of fifteen other vaccines based on this technology, including Qaz- COVID-in (NCT04691908), BBV152 (NCT04641481), BBIBP-CorV (NCT04560881, NCT04510207), COVIran-Barekat (IRCT20201202049567N1, IRCT20201202049567N2), and Sinovac Biotech Ltd’s CoronaVac which is currently used in UAE and China [111].
Replicating and non-replicating viral vectors
Vaccines based on viral vectors have several benefits such as releasing antigens in a longer period of time, high antigenic protein expression and provoking stronger immune responses. Moreover, compared to recombinant proteins and inactivated viruses, this platform benefits from having the capability to stimulate more immunoglobulin A release, cause stronger mucosal immunity and consequently increase the strength of barriers’ resistance to virus entry [11]. Despite its notable advantages, this platform is considered to be of less efficacy due to previously existing immunity against the utilized viral strain used as the vector [112], however, this limitation can often be partially compensated by priming this vaccine with a different vaccine or by adding a booster dose to the vaccination protocol [113, 114]. A vaccine based on recombinant and non-replicating adenoviral vectors, mostly known as Ad5-nCoV, has been designed by the CanSino Biological Inc. company and is undergoing phase III (NCT04526990) of its clinical trials [115,116,117]. In total, there are two replicating and seventeen non-replicating viral vector-based vaccines that are now being investigated in clinical stages [111].
DNA based vaccines
This category of SARS-CoV-2 vaccines mainly transfers the S gene of this virus into certain bacterial plasmids including CMVs [118]. Having the benefit of stimulating fast industrial production as well as being highly flexible toward antigenic domain manipulations and targeting both the humoral and cellular immune systems, DNA-based vaccines are considered to be promising platforms for vaccine development [119]. There are now eleven DNA-based vaccine candidates in for SARS-CoV-2, three of which are now passing phase III of their clinical examinations [111].
RNA based vaccines
Similar to DNA-based vaccines, this group of vaccines benefit from significant flexibility for antigenic domains changes [119, 120]. After less than three months from the publication of the SARS-CoV-2 sequence, Moderna Company succeeded in taking their vaccine into the clinical phases. This vaccine is an RNA-based (mRNA-1273) vaccination platform that stimulates the immune system against SARS-CoV-2 spike protein and is delivered into cells via lipid nanostructures [121]. BioNTech, Fosun Pharma, and Pfizer companies have also developed vaccines based on this technology that are currently being exploited worldwide. In total, 18 RNA-based vaccines have been designed so far that are now going through clinical trial studies.
Live attenuated viral vaccines
Being mainly produced by the use of live weakened viral strains or non-virulent species, this category of vaccines are capable of stimulating significant mucosal and cellular responses, similar to those caused by the natural invasion of the virus. Despite the employment of efficient methods in order to attenuate viruses (such as storage in low temperatures and genetic modification) [122, 123], SARS-CoV-2 vaccines built upon this technology suffer from several drawbacks, including high risks of unintentional viral transmission to the unvaccinated population through feces, existing possibility of viral species recombination in the host’s body, as well as difficult formulation and quality control processes that decelerate the large-scale industrial production of vaccine [124].
This group of vaccines have the advantage of inducing significant immunization and also triggering both humoral and cellular immunity concurrently. Different viral antigens are capable of triggering the immune system thus resulting in the generation of a diverse range of antibodies and T memory cells. There are currently two vaccines with this platform (COVI-VAC (NCT04619628) and MV-014-212 (NCT04798001)) that have recently moved to clinical trial stages [111].
Protein subunit-based vaccines
Due to their desirably safe nature and efficient humoral immunity stimulation, protein subunit vaccines are the most abundant group of SARS-CoV-2 vaccines. The main mechanism of action of these vaccines mostly relies on the expression of the Spike glycoprotein or even just the Spike’s RBD region [125]. The expression of the Spike protein is associated with moderate difficulty and has thus raised concerns regarding the production of vaccines with the use of this technology. However, several methods such as exclusive expression of RBD, have been proposed for minimizing this drawback [11]. There are now thirty-eight vaccine candidates that reside in this category, twelve of which are currently in Phase II/III and III of clinical trials. Novavax (NCT04611802), FINLAY-FR-2 (RPCEC00000354), EpiVacCorona (NCT04780035), and Vaxxinity (NCT04683224) are some of the examples of vaccines that have been developed with this technology Table 3 [111].
Animal models for the evaluation of SARS-CoV-2 vaccination efficacy
The utilization of animal models for testing the efficacy of vaccines is a common yet critically important subject. In the case of SARS, high specificity of this virus’s S protein to human ACE2 receptors had previously made it difficult to find a proper animal model for testing vaccination efficacy in the past, but with creating transgenic mice that express human ACE2 receptor proteins this issue was mainly solved. Due to the similarity of ACE2 receptors in both SARS and COVID-19 infections, those transgenic mice are likely to be suitable for use for COVID-19 as well. Rhesus macaques have also been successfully used for testing COVID-19 vaccines by Sinovac companies in China [147, 148]. For SARS and MERS, other animals such as rabbits, hamsters, ferrets, pigs, marmosets, cats, and cows have been used as animal models [149, 150].
Vaccination efficacy against the Delta variant
Recent literature has displayed that vaccine-induced immunity followed by one dose of BNT162b2 or ChAdOx1 nCoV-19 vaccines was about 48.7% and 30.7% against the Alpha and Delta variants respectively. Vaccine-induced immunity with two doses of BNT162b2 was also reported to be about 93.7% and 88.0% against the Alpha and Delta viral variants while being 74.5% and 67% for the ChAdOx1 nCoV-19 vaccine, all in a respective order [9]. Similarly, another investigation has recently reported that this new Delta variant possesses 6.8 times higher resistance to sera-induced neutralization in individuals who were vaccinated with either Moderna or Pfizer vaccines. Accordingly, neutralizing antibody titer in infected and vaccinated people followed by exposure with the Delta variant was shown to have declined dramatically, nevertheless, 79% of all samples of sera obtained from mRNA vaccinated individuals were able to neutralize the Delta viral variant [110]. While information regarding other vaccines is limited, a study constructed in Scotland investigated the vaccination efficacies of ChAdOx1 nCoV-19 (AstraZeneca) and BNT162b2 (Pfizer–BioNTech vaccine) against the B.1.617.2 SARS-CoV-2 variant. Both of these vaccines were reported to reduce infection risks and hospitalization post infection with Delta variant, however, they were less efficient compared to their immunity against the Alpha variant, which is consistent with previous results [151].
Of note, obtained viral loads in nasal swabs of vaccinated and unvaccinated individuals were reported to be similarly followed by exposure with the Delta variant. Moreover, 68% of the infected individuals who had been vaccinated beforehand were tested PCR-positive for COVID-19 infection, even though 8 of them did not suffer from symptoms of infection at the time of PCR. These findings were indicative of the importance of wearing masks despite the state of vaccination, as it appears that vaccinated individuals might also be capable of spreading the virus [152]. Lastly, due to the limited data addressing the efficacy of vaccines against the Delta variant of COVID-19, there is still a lot of doubt revolving around this matter. However, currently available data confirms the crucial necessity for vaccination, as it can still provide the public with lower yet excellent immunity against severe SARS-CoV-2 infection.
Challenges in vaccine development regulatory systems
Vaccine development with passing phases I, II, and III of clinical trials and getting approval licensures is a rather long process that normally requires 6 to 11 years [153]. Before clinical trials, regulatory agencies should check production processes and information obtained from preclinical tests for vaccine safety assurance [154].
Even though a lot of attention is directed towards global candidate vaccine efficacy, it should be noted that even if a vaccine is capable of reducing disease severity but not preventing it completely, it’s still a potential candidate for decreasing mortality rates significantly [155]. Therefore, while some vaccines might lack the desired and ideal efficiency, they can still contribute greatly to viral shedding and preventing the rapid spread of the virus [11, 156]. Due to the relevantly high risk for vaccine failure with the emergence of new variants, vaccine investors often define multiple steps for the manufacturing and the process is paused after each step in order to make sure of chances of success for that part, and then further investments take place for the next steps [157]. It is clear that this method for vaccine development takes a lot of time and is likely to be inefficient for a fast pandemic or epidemic situation, as a large population will probably be infected by the time that vaccine is fully developed. It has been suggested that for speeding up this process, industrial production of vaccines with higher chances of success and efficacy (according to information from phases I and II of clinical trials) should be initiated so they will be available for the public to use soon after receiving efficacy approval [153, 158]. Even though obtaining and mass production of a vaccine isn’t a limiting step, but multi-step production and obtainment of approvals from regulatory agencies turn this into a rather long and time-consuming process. The occurrence of several epidemics in the past two decades has challenged the efficiency of this time-consuming process in protecting public health. It seems that a change of routine, methods, and regulatory agencies’ rules for faster scrutiny of safety measures, effectiveness and possible side effects of vaccines and subsequently developing faster approaches and methods for vaccine evaluation is necessary [159, 160].
Conclusion
With the continuous evolution of SARS-CoV-2, the emergence of novel and potentially more fatal variants is now a well-established phenomenon. Mutations within the genome and amino acid profiles of SARS-CoV-2 are affected by several factors, and while some of the observed mutations cease to exist in the next generations of SARS-CoV-2, there are several mutations that are passed on to the next mutated strains and further enhance their pathogenicity and transmissibility through different pathways and mechanisms. Aside from the increased infection severity and faster spread, the impact of mutations on vaccination efficacy is now one of the most serious concerns regarding this matter. Being mainly constructed against certain previous VOCs, it is now unclear whether the developed vaccines are capable of providing sufficient immunity toward the newly emerging strains of SARS-CoV-2. With such doubts in mind, recent studies of vaccines have been mainly indicative of reduced but yet still desirable immunity against new VOCs, and further confirming the fact that global vaccination is still a reliable platform for overcoming this battle of humans against coronaviruses. With that in mind, this study was constructed with the aim of exploring recent advances regarding SARS-CoV-2 mutations as well as providing readers with comprehensive information about some of the latest COVID-19 vaccines and aid them in gaining insight regarding where we now stand in this crisis, and what to expect.
References
Wang W, Xu Y, Gao R, Lu R, Han K, Wu G, et al. Detection of SARS-CoV-2 in different types of clinical specimens. JAMA - J Am Med Assoc. 2020;323(18):1843–4.
Davies NG, Abbott S, Barnard RC, Jarvis CI, Kucharski AJ, Munday JD, et al. Estimated transmissibility and impact of SARS-CoV-2 lineage B.1.1.7 in England. Science (80-). 2021;372(6538):eabg3055.
Adam D. What scientists know about new, fast-spreading coronavirus variants. Nature. 2021;594(7861):19–20.
Fehr AR, Perlman S, Fehr AR, Perlman S. Coronaviruses: an overview of their replication and pathogenesis. Coronaviruses Methods Protoc. 2015;1282(1):1–23.
Harvey WT, Carabelli AM, Jackson B, Gupta RK, Thomson EC, Harrison EM, et al. SARS-CoV-2 variants, spike mutations and immune escape. Nat Rev Microbiol. 2021;19(7):409–24.
Banoun H. Evolution of SARS-CoV-2: review of mutations, role of the host immune system. Nephron. 2021;145(4):392–403.
Na W, Moon H, Song D. A comprehensive review of SARS-CoV-2 genetic mutations and lessons from animal coronavirus recombination in one health perspective. J Microbiol. 2021;59(3):332–40.
Edara VV, Norwood C, Floyd K, Lai L, Davis-Gardner ME, Hudson WH, et al. Infection- and vaccine-induced antibody binding and neutralization of the B.1.351 SARS-CoV-2 variant. Cell Host Microbe. 2021;29(4):516–521.e3.
Lopez Bernal J, Andrews N, Gower C, Gallagher E, Simmons R, Thelwall S, et al. Effectiveness of Covid-19 Vaccines against the B.1.617.2 (Delta) Variant. N Engl J Med. 2021.
Plotkin S, Robinson JM, Cunningham G, Iqbal R, Larsen S. The complexity and cost of vaccine manufacturing – an overview. Vaccine. 2017;35(33):4064–71.
Krammer F. SARS-CoV-2 vaccines in development. Nature. 2020;586(7830):516–27.
Chakraborty S, Mallajosyula V, Tato CM, Tan GS, Wang TT. SARS-CoV-2 vaccines in advanced clinical trials: where do we stand? Adv Drug Deliv Rev. 2021;172:314–38. Available from: https://reader.elsevier.com/reader/sd/pii/S0169409X21000247?token=37A0293B89855BF59F5D6BB5192A253719943386B4679E9E542CF7EB0865CA9A11F97F7ACD7AE717F406184D4681213B&originRegion=eu-west-1&originCreation=20210602145950. Accessed 20 Jan 2021.
Du Y, Tu L, Zhu P, Mu M, Wang R, Yang P, et al. Clinical features of 85 fatal cases of COVID-19 from Wuhan: a retrospective observational study. Am J Respir Crit Care Med. 2020;201(11):1372–9.
Abbasinia M, Hormati A, Eshagh Hossaini SK, Foroghi Ghomi SY, Zamani F, Afifian M, et al. Clinical manifestations of gastrointestinal symptoms in COVID-19 patients: an integrative review. Gastroenterol Nurs. 2021;44(1):E1–10.
Robba C, Battaglini D, Pelosi P, Rocco PRM. Multiple organ dysfunction in SARS-CoV-2: MODS-CoV-2. Expert Rev Respir Med. 2020;14(9):865–8.
Ramírez P, Gordón M, Martín-Cerezuela M, Villarreal E, Sancho E, Padrós M, et al. Acute respiratory distress syndrome due to COVID-19. Clinical and prognostic features from a medical critical care unit in Valencia, Spain. Med Int. 2021;45(1):27–34.
Ufuk F, Savaş R. Chest CT features of the novel coronavirus disease (COVID-19). Turkish J Med Sci. 2020;50(4):664–78.
Pan F, Ye T, Sun P, Gui S, Liang B, Li L, et al. Time course of lung changes at chest CT during recovery from coronavirus disease 2019 (COVID-19). Radiology. 2020;295(3):715–21.
Clark R, Waters B, Stanfill AG. Elevated liver function tests in COVID-19. Nurs Pract. 2021;46(1):21–6.
Dehghanbanadaki H, Aazami H, Shabani M, Amighi D, Seif F, Zare Dehnavi A, et al. A systematic review and meta-analysis on the association between lymphocyte subsets and the severity of COVID-19. Immunopathol Persa 2022;x(x). https://doi.org/10.34172/ipp.2022.xx
Nishiga M, Wang DW, Han Y, Lewis DB, Wu JC. COVID-19 and cardiovascular disease: from basic mechanisms to clinical perspectives. Nat Rev Cardiol. 2020;17(9):543–58.
Hosseinkhani S, Arjmand B, Bandarian F, Aazami H, Hadizadeh N, Najjar N, et al. Omics experiments in Iran, a review in endocrine and metabolism disorders studies. J Diabetes Metab Disorders. 2021.
Steenblock C, Schwarz PEH, Ludwig B, Linkermann A, Zimmet P, Kulebyakin K, et al. COVID-19 and metabolic disease: mechanisms and clinical management. Lancet Diabetes Endocrinol. 2021;9(11):786–98.
Heaney AI, Griffin GD, Simon EL. Newly diagnosed diabetes and diabetic ketoacidosis precipitated by COVID-19 infection. Am J Emerg Med. 2020;38(11):2491.e3–4.
Montefusco L, Ben Nasr M, D’Addio F, Loretelli C, Rossi A, Pastore I, et al. Acute and long-term disruption of glycometabolic control after SARS-CoV-2 infection. Nat Metab. 2021;3(6):774–85.
Mousavi T, Hadizadeh N, Nikfar S, Abdollahi M. Drug discovery strategies for modulating oxidative stress in gastrointestinal disorders. Expert Opin Drug Discovery. 2020;15(11):1309–41.
Cecchini R, Cecchini AL. SARS-CoV-2 infection pathogenesis is related to oxidative stress as a response to aggression. Med Hypotheses. 2020;143:110102.
Maestre-Muñiz MM, Arias Á, Mata-Vázquez E, Martín-Toledano M, López-Larramona G, Ruiz-Chicote AM, et al. Long-term outcomes of patients with coronavirus disease 2019 at one year after hospital discharge. J Clin Med. 2021;10(13):2945.
Kumaran NK, Karmakar BK, Taylor OM. Coronavirus disease-19 (COVID-19) associated with acute necrotising pancreatitis (ANP). BMJ Case Rep. 2020;13(9):e237903.
Kanji JN, Zelyas N, MacDonald C, Pabbaraju K, Khan MN, Prasad A, et al. False negative rate of COVID-19 PCR testing: a discordant testing analysis. Virol J. 2021;18(1):1–6.
Vandenberg O, Martiny D, Rochas O, van Belkum A, Kozlakidis Z. Considerations for diagnostic COVID-19 tests. Nat Rev Microbiol. 2021;19(3):171–83.
Judson SD, Munster VJ. Nosocomial transmission of emerging viruses via aerosol-generating medical procedures. Viruses. 2019;11(10):940.
Patrucco F, Albera C, Bellocchia M, Foci V, Gavelli F, Castello LM, et al. SARS-CoV-2 detection on Bronchoalveolar lavage: an Italian multicenter experience. Respiration. 2021;99(11):970–8.
Shokraee K, Mahdavi H, Panahi P, Seirafianpour F, Jahromizadeh AM, Tofighi R, et al. Accuracy of chest computed tomography and reverse transcription polymerase chain reaction in diagnosis of 2019 novel coronavirus disease; a systematic review and meta-analysis. Immunopathol Persa. 2021;7(2):e36.
Huybens EM, Bus MPA, Massaad RA, Wijers L, van der Voet JA, Delfos NM, et al. Correlation of chest CT and RT-PCR testing for coronavirus disease 2019 (COVID-19) in China: a report of 1014 cases. Radiology. 2020;296(2):1–2. Available from. https://doi.org/10.1016/j.oraloncology.2020.105043.
Yadav SK, Akhter Y. Statistical modeling for the prediction of infectious disease dissemination with special reference to COVID-19 spread. Front Public Health. 2021;9:680.
Lauer SA, Grantz KH, Bi Q, Jones FK, Zheng Q, Meredith HR, et al. The incubation period of coronavirus disease 2019 (CoVID-19) from publicly reported confirmed cases: estimation and application. Ann Intern Med. 2020;172(9):577–82.
Backer JA, Klinkenberg D, Wallinga J. Incubation period of 2019 novel coronavirus (2019- nCoV) infections among travellers from Wuhan, China, 20 28 January 2020. Eurosurveillance. 2020;25(5):2000062.
Bowser AD. Some Patients Could Show COVID-19 Symptoms After Quarantine [Internet]. 2020. Available from: https://www.medscape.com/viewarticle/926507. Accessed 9 Mar 2020
Eshrati B, Baradaran HR, Moradi G, Dehghanbanadaki H, Azh N, Soheili M, et al. Evaluation of reinfection in COVID-19 patients in the world: a narrative review. Med J Islam Repub Iran. 2021;35(1):1–13.
Walker PJ, Siddell SG, Lefkowitz EJ, Mushegian AR, Dempsey DM, Dutilh BE, et al. Changes to virus taxonomy and the international code of virus classification and nomenclature ratified by the international committee on taxonomy of viruses (2019). Arch Virol. 2019;164(9):2417–29.
Helmy YA, Fawzy M, Elaswad A, Sobieh A, Kenney SP, Shehata AA. The COVID-19 pandemic: a comprehensive review of taxonomy, genetics, epidemiology, diagnosis, treatment, and control. J Clin Med. 2020;9(4):1225.
Gorbalenya AE, Krupovic M, Mushegian A, Kropinski AM, Siddell SG, Varsani A, et al. The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks. Nat Microbiol. 2020;5(5):668–74.
Volz E, Mishra S, Chand M, Barrett JC, Johnson R, Geidelberg L, et al. Assessing transmissibility of SARS-CoV-2 lineage B.1.1.7 in England. Nature. 2021;593(7858):266–9.
European Centre for Disease Prevention and Control. SARS-CoV-2-increased cir culation of variants of concern and vaccine rollout in the EU/EEA. [Internet]. Available from: https://www.brief.com.cy/sites/default/files/2021-03/RRA-covid-19-14th update-15-feb-2021.pd.
Graham MS, Sudre CH, May A, Antonelli M, Murray B, Varsavsky T, et al. Changes in symptomatology, reinfection, and transmissibility associated with the SARS-CoV-2 variant B.1.1.7: an ecological study. Lancet Public Health. 2021;6(5):e335–45.
Faria NR, Claro IM, Candido D, Franco LAM, Andrade PS, Thais M, et al. Genomic characterisation of an emergent SARS-CoV-2 lineage in Manaus: preliminary findings. VirologicalOrg [Internet]. 2021;1–9. Available from: https://virological.org/t/genomic-characterisation-of-an-emergent-sars-cov-2-lineage-in-manaus-preliminary-findings/586. Accessed 14 Apr 2021.
Rambaut A, Loman N, Pybus O, Barclay W, Barrett J, Carabelli A, et al. Preliminary genomic characterisation of an emergent SARS-CoV-2 lineage in the UK defined by a novel set of spike mutations - SARS-CoV-2 coronavirus / nCoV-2019 Genomic Epidemiology - Virological. Virological.org [Internet]. 2020;1–9. Available from: https://virological.org/t/preliminary-genomic-characterisation-of-an-emergent-sars-cov-2-lineage-in-the-uk-defined-by-a-novel-set-of-spike-mutations/563. Accessed 19 Dec 2020.
Tegally H, Wilkinson E, Giovanetti M, Iranzadeh A, Fonseca V, Giandhari J, et al. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv. 2020.
Campbell F, Archer B, Laurenson-Schafer H, Jinnai Y, Konings F, Batra N, et al. Increased transmissibility and global spread of SARSCoV- 2 variants of concern as at June 2021. Eurosurveillance. 2021;26(24):1–6.
Faria NR, Mellan TA, Whittaker C, Claro IM, Candido DDS, Mishra S, et al. Genomics and epidemiology of the P.1 SARS-CoV-2 lineage in Manaus, Brazil. Science (80-). 2021;372:6544.
Lucaci A, Zehr J, Shank S, Bouvier D, Mei H, Nekrutenko A, et al. RASCL: Rapid assessment of SARS-COV-2 clades enabled through molecular sequence analysis and its application to B.1.617.1 and B.1.617.2 [Internet]. Virological.Org. 2021. Available from: https://virological.org/t/rascl-rapid-assessment-of-sars-cov-2-clades-enabled-through-molecular-sequence-analysis-and-its-application-to-b-1-617-1-and-b-1-617-2/709. Accessed 29 May 2021.
Diseases NI for C. Frequently asked questions for the B.1.1.529 mutated SARS-CoV-2 lineage in South Africa. 2021;
Centers for Disease Control and Prevention. Science Brief: Omicron (B.1.1.529) Variant [Internet]. CDC COVID-19 Science Briefs. 2020. Available from: http://www.ncbi.nlm.nih.gov/pubmed/34932278. Accessed 2 Dec 2021.
WHO. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern [Internet]. Who. 2021. p. 1. Available from: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern. Accessed 15 Jan 2021.
Pearson CA, Russell TW, Davies NG, Kucharski AJ, CMMID COVID-19 working group, Edmunds WJ, et al. Estimates of severity and transmissibility of novel South Africa SARS-CoV-2 variant 501Y.V2. Preprint. 2021;50:1–4 Available from: https://cran.r-project.org/package=wpp2019%0Ahttps://cmmid.github.io/topics/covid19/sa-novel-variant.html. Accessed 11 Dec 2021.
Buss LF, Prete CA, Abrahim CMM, Mendrone A, Salomon T, De Almeida-Neto C, et al. Three-quarters attack rate of SARS-CoV-2 in the Brazilian Amazon during a largely unmitigated epidemic. Science (80-). 2021;371(6526):288–92.
Jaimes JA, André NM, Chappie JS, Millet JK, Whittaker GR. Phylogenetic analysis and structural modeling of SARS-CoV-2 spike protein reveals an evolutionary distinct and Proteolytically sensitive activation loop. J Mol Biol. 2020;432(10):3309–25.
Wang H, Li X, Li T, Zhang S, Wang L, Wu X, et al. The genetic sequence, origin, and diagnosis of SARS-CoV-2. Eur J Clin Microbiol Infect Dis. 2020;39(9):1629–35.
Boni MF, Lemey P, Jiang X, Lam TTY, Perry B, Castoe T, et al. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. bioRxiv. 2020.
Jungreis I, Sealfon R, Kellis M. SARS-CoV-2 gene content and COVID-19 mutation impact by comparing 44 Sarbecovirus genomes. Nat Commun. 2021;12(1):1–20.
Nguyen TT, Pathirana PN, Nguyen T, Nguyen QVH, Bhatti A, Nguyen DC, et al. Genomic mutations and changes in protein secondary structure and solvent accessibility of SARS-CoV-2 (COVID-19 virus). Sci Rep. 2021;11(1):3487.
Mercatelli D, Giorgi FM. Geographic and genomic distribution of SARS-CoV-2 mutations. Front Microbiol. 2020;11:1800.
Roujian Lu, Xiang Zhao M, Juan Li P, Peihua Niu P, Bo Yang, Honglong Wu, et al. Genomic characterisation and epidemiology of 2019 novel coronavirus: implications for virus origins and receptor binding. Lancet [Internet]. 2020; Available from: https://www.thelancet.com/article/S0140-6736(20)30251-8/fulltext. Accessed 30 Jan 2020.
Naqvi AAT, Fatima K, Mohammad T, Fatima U, Singh IK, Singh A, et al. Insights into SARS-CoV-2 genome, structure, evolution, pathogenesis and therapies: structural genomics approach. Biochim Biophys Acta Mol basis Dis. 2020;1866(10):165878.
Dagur HS. Genome Organization of Covid-19 and emerging severe acute respiratory syndrome Covid-19 outbreak: a pandemic. Eurasian. J Med Oncol. 2020.
Gordon DE et al. A SARS-CoV-2 Protein Interaction Map Reveals Targets for Drug-Repurposing. Gordon et al (Nevan). Nature Ap2020 FINAL. Nature. 2020;583:459–68. Available from: https://www.nature.com/articles/s41586-020-2286-9. Accessed 23 Apr 2021.
Min YQ, Mo Q, Wang J, Deng F, Wang H, Ning YJ. SARS-CoV-2 nsp1: bioinformatics, potential structural and functional features, and implications for drug/vaccine designs. Front Microbiol. 2020;11:29.
Schubert K, Karousis ED, Jomaa A, Scaiola A, Echeverria B, Gurzeler LA, et al. SARS-CoV-2 Nsp1 binds the ribosomal mRNA channel to inhibit translation. Nat Struct Mol Biol. 2020;27(10):959–66.
Xia H, Cao Z, Xie X, Zhang X, Chen JYC, Wang H, et al. Evasion of type I interferon by SARS-CoV-2. Cell Rep. 2020;33(1):108234.
Yadav R, Chaudhary JK, Jain N, Chaudhary PK, Khanra S, Dhamija P, et al. Role of structural and non-structural proteins and therapeutic targets of SARS-CoV-2 for COVID-19. Cells. 2021;10(4):821.
Nelson CW, Ardern Z, Goldberg TL, Meng C, Kuo CH, Ludwig C, et al. Dynamically evolving novel overlapping gene as a factor in the SARS-CoV-2 pandemic. Elife. 2020;9:1–45.
Casalino L, Gaieb Z, Goldsmith JA, Hjorth CK, Dommer AC, Harbison AM, et al. Beyond shielding: the roles of glycans in the SARS-CoV-2 spike protein. ACS Cent Sci. 2020;6(10):1722–34.
Chan JFW, Kok KH, Zhu Z, Chu H, To KKW, Yuan S, et al. Genomic characterization of the 2019 novel human-pathogenic coronavirus isolated from a patient with atypical pneumonia after visiting Wuhan. Emerg Microbes Infect. 2020;9(1):221–36.
Mohammadi E, Shafiee F, Shahzamani K, Ranjbar MM, Alibakhshi A, Ahangarzadeh S, et al. Novel and emerging mutations of SARS-CoV-2: biomedical implications. Biomed Pharmacother. 2021;139:111599.
Wang L, Xiang Y. Spike glycoprotein-mediated entry of sars coronaviruses. Viruses. 2020;12(11):1289.
Ou X, Liu Y, Lei X, Li P, Mi D, Ren L, et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat Commun. 2020;11(1):1–12.
Zhang Q, Xiang R, Huo S, Zhou Y, Jiang S, Wang Q, et al. Molecular mechanism of interaction between SARS-CoV-2 and host cells and interventional therapy. Signal Transduct Target Ther. 2021;6(1):1–19.
Morse JS, Lalonde T, Xu S, Liu WR. Learning from the past: possible urgent prevention and treatment options for severe acute respiratory infections caused by 2019-nCoV. ChemBioChem. 2020;21(5):730–8.
Robson F, Khan KS, Le TK, Paris C, Demirbag S, Barfuss P, et al. Coronavirus RNA proofreading: molecular basis and therapeutic targeting. Mol Cell. 2020;80(6):1136–8.
Ong SYQ, Flyamer IM, Bickmore WA, Biddie SC. From bedside to bench: regulation of host factors in SARS-CoV-2 infection. Exp Mol Med. 2021;53(4):483–94.
Zhang XX, Tan Y, Ling Y, Lu G, Liu F, Yi Z, et al. Viral and host factors related to the clinical outcome of COVID-19. Nature. 2020;583(7816):437–40.
Yang X, Dong N, Chan EWC, Chen S. Genetic cluster analysis of SARS-CoV-2 and the identification of those responsible for the major outbreaks in various countries. Emerg Microbes Infect. 2020;9(1):1287–99.
Yang HC, Chen CWCH, Wang JH, Liao HC, Yang CT, Chen CWCH, et al. Analysis of genomic distributions of SARS-CoV-2 reveals a dominant strain type with strong allelic associations. Proc Natl Acad Sci U S A. 2020;117(48):30679–86.
Zhao Z, Sokhansanj BA, Malhotra C, Zheng K, Rosen GL. Genetic grouping of SARS-CoV-2 coronavirus sequences using informative subtype markers for pandemic spread visualization. bioRxiv. 2020;16(9):e1008269.
Xiaolu T, Changcheng W, Xiang L, Yuhe S, Xinmin Y, Xinkai W, et al. On the origin and continuing evolution of SARS-CoV-2. Natl Sci Rev. 2020;7(06):1012–23 Available from: https://academic.oup.com/nsr/article-abstract/7/6/1012/5775463. Accessed 3 Mar 2020.
Ruan Y, Luo Z, Tang X, Li G, Wen H, He X, et al. On the founder effect in COVID-19 outbreaks: how many infected travelers may have started them all? Natl Sci Rev. 2021;8(1).
Andreano E, Piccini G, Licastro D, Casalino L, Johnson N V, Paciello I, et al. SARS-CoV-2 escape in vitro from a highly neutralizing COVID-19 convalescent plasma. bioRxiv Prepr Serv Biol [Internet]. 2020.
Jia Z, Gong W. Will mutations in the spike protein of SARS-CoV-2 Lead to the failure of COVID-19 vaccines? J Korean Med Sci. 2021;26(18):1–11.
Zinzula L. Lost in deletion: the enigmatic ORF8 protein of SARS-CoV-2. Biochem Biophys Res Commun. 2021;538:116–24.
Pereira F. Evolutionary dynamics of the SARS-CoV-2 ORF8 accessory gene. Infect Genet Evol. 2020;85:104525.
Resende P SARS-CoV-2 reinfection by the new Variant of Concern (VOC) P.1 in Amazonas, Brazil - SARS-CoV-2 coronavirus / nCoV-2019 Genomic Epidemiology - Virological. Virological [Internet]. 2021;(February 2020):1–9. Available from: https://virological.org/t/sars-cov-2-reinfection-by-the-new-variant-of-concern-voc-p-1-in-amazonas-brazil/596. Accessed 18 Jan 2021.
Liu L, Wang P, Nair MS, Yu J, Rapp M, Wang Q, et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature. 2020;584(7821):450–6.
McCallum M, De Marco A, Lempp FA, Tortorici MA, Pinto D, Walls AC, et al. N-terminal domain antigenic mapping reveals a site of vulnerability for SARS-CoV-2. Cell. 2021;184(9):2332–2347.e16.
Suryadevara N, Shrihari S, Gilchuk P, VanBlargan LA, Binshtein E, Zost SJ, et al. Neutralizing and protective human monoclonal antibodies recognizing the N-terminal domain of the SARS-CoV-2 spike protein. Cell. 2021;184(9):2316–2331.e15.
Greaney AJ, Loes AN, Crawford KHD, Starr TN, Malone KD, Chu HY, et al. Comprehensive mapping of mutations in the SARS-CoV-2 receptor-binding domain that affect recognition by polyclonal human plasma antibodies. Cell Host Microbe. 2021;29(3):463–476.e6.
Duval R, Moretti TMT, Morettti MT, Representations F, Maggio DP, Soares MAS, et al. Escape from neutralizing antibodies by SARS-CoV-2 spike protein variants 2. Revemat Rev Eletrôn Educ Matemática. 2018;13(1):1689–99.
Baum A, Fulton BO, Wloga E, Copin R, Pascal KE, Russo V, et al. Antibody cocktail to SARS-CoV-2 spike protein prevents rapid mutational escape seen with individual antibodies. Science (80- ). 2020;369(6506):1014–8.
Thomson EC, Rosen LE, Shepherd JG, Spreafico R, da Silva FA, Wojcechowskyj JA, et al. Circulating SARS-CoV-2 spike N439K variants maintain fitness while evading antibody-mediated immunity. Cell. 2021;184(5):1171–1187.e20.
Chi X, Yan R, Zhang J, Zhang G, Zhang Y, Hao M, et al. A neutralizing human antibody binds to the N-terminal domain of the spike protein of SARS-CoV-2. Science (80-). 2020;369(6504):650–5.
McCarthy KR, Rennick LJ, Nambulli S, Robinson-McCarthy LR, Bain WG, Haidar G, et al. Recurrent deletions in the SARS-CoV-2 spike glycoprotein drive antibody escape. Science (80-). 2021;371(6534):1139–42.
Hodcroft EB, Zuber M, Nadeau S, Crawford KHD, Bloom JD, Veesler D, et al. Emergence and spread of a SARS-CoV-2 variant through Europe in the summer of 2020. medRxiv Prepr Serv Heal Sci [Internet]. 2020; Available from: http://www.ncbi.nlm.nih.gov/pubmed/33269368; http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=PMC7709189. Accessed 24 Mar 2020.
Nextstrain, Nextrain. Genomic epidemiology of novel coronavirus - global subsampling. Nextstrain [Internet]. 2020;1.
Garcia-Beltran WF, Lam EC, St. Denis K, Nitido AD, Garcia ZH, Hauser BM, et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell. 2021;184(9):2523.
Garcia-Beltran WF, St Denis KJ, Hoelzemer A, Lam EC, Nitido AD, Sheehan ML, et al. mRNA-based COVID-19 vaccine boosters induce neutralizing immunity against SARS-CoV-2 Omicron variant. medRxiv Prepr Serv Heal Sci [Internet]. 2021.
Cele S, Jackson L, Khan K, Khoury D, Moyo-Gwete T, Tegally H, et al. SARS-CoV-2 Omicron has extensive but incomplete escape of Pfizer BNT162b2 elicited neutralization and requires ACE2 for infection. Mahomed-Yunus S Moosa [Internet]. 10:11. Available from: https://covdb.stanford.edu/page/mutation-viewer/#sec_b-1-351. Accessed 17 Dec 2021.
Yazdani M, Khezri J, Hadizadeh N, Amir Zakaria J, Naderi M, Mahmoodian S, et al. Depinar, a drug that potentially inhibits the binding and entry of COVID-19 into host cells based on computer-aided studies. Res Pharm Sci. 2021;16(3):315–25.
Yadav M, Dhagat S, Eswari JS. Emerging strategies on in silico drug development against COVID-19: challenges and opportunities. Eur J Pharm Sci. 2020;155:105522.
Mei M, Tan X. Current strategies of antiviral drug discovery for COVID-19. Front Mol Biosci. 2021;8:310.
Edara V-V, Pinsky BA, Suthar MS, Lai L, Davis-Gardner ME, Floyd K, et al. Infection and vaccine-induced neutralizing-antibody responses to the SARS-CoV-2 B.1.617 variants. N Engl J Med. 2021;385(7):664–6 Available from: https://www.nejm.org/doi/full/10.1056/NEJMc2107799. Accessed 12 Aug 2021.
WHO. COVID-19 vaccine tracker and landscape [Internet]. Who. 2021. Available from: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines
Nayak S, Herzog RW. Progress and prospects: immune responses to viral vectors. Gene Ther. 2010;17(3):295–304.
Yang ZY, Wyatt LS, Kong WP, Moodie Z, Moss B, Nabel GJ. Overcoming immunity to a viral vaccine by DNA priming before vector boosting. J Virol. 2003;77(1):799–803 Available from: http://ovidsp.ovid.com/ovidweb.cgi?T=JS&PAGE=reference&D=emed6&NEWS=N&AN=2003001409. Accessed January 2003.
Pandey A, Singh N, Vemula SV, Couëtil L, Katz JM, Donis R, et al. Impact of preexisting adenovirus vector immunity on immunogenicity and protection conferred with an adenovirus-based H5N1 influenza vaccine. PLoS One. 2012;7(3):e33428.
Zhu FC, Li YH, Guan XH, Hou LH, Wang WJ, Li JX, et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: a dose-escalation, open-label, non-randomised, first-in-human trial. Lancet. 2020;395(10240):1845–54.
Zhu FC, Guan XH, Li YH, Huang JY, Jiang T, Hou LH, et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: a randomised, double-blind, placebo-controlled, phase 2 trial. Lancet. 1024.
Inc. CB and BI of B. Phase III Trial of A COVID-19 Vaccine of Adenovirus Vector in Adults 18 Years Old and Above. ClinicalTrials.gov [Internet]. 2020; Available from: https://clinicaltrials.gov/ct2/show/NCT04526990. Accessed 22 Jan 2021.
Nagy A, Alhatlani B. An overview of current COVID-19 vaccine platforms. Comput Struct Biotechnol J. 2021;19:2508–17.
Thanh Le T, Andreadakis Z, Kumar A, Gómez Román R, Tollefsen S, Saville M, et al. The COVID-19 vaccine development landscape. Nat Rev Drug Discov. 2020;19(5):305–6.
Maruggi G, Zhang C, Li J, Ulmer JB, Yu D. mRNA as a transformative Technology for Vaccine Development to control infectious diseases. Mol Ther. 2019;27(4):757–72.
Wu K, Werner AP, Koch M, Choi A, Narayanan E, Stewart-Jones GBE, et al. Serum neutralizing activity elicited by mRNA-1273 vaccine. N Engl J Med. 2021;384(15):1468–70.
Talon J, Salvatore M, O’Neill RE, Nakaya Y, Zheng H, Muster T, et al. Influenza a and B viruses expressing altered NS1 proteins: a vaccine approach. Proc Natl Acad Sci U S A. 2000;97(8):4309–14.
Broadbent AJ, Santos CP, Anafu A, Wimmer E, Mueller S, Subbarao K. Evaluation of the attenuation, immunogenicity, and efficacy of a live virus vaccine generated by codon-pair bias de-optimization of the 2009 pandemic H1N1 influenza virus, in ferrets. Vaccine. 2016;34(4):563–70.
Frederiksen LSF, Zhang Y, Foged C, Thakur A. The long road toward COVID-19 herd immunity: vaccine platform technologies and mass immunization strategies. Front Immunol. 2020;11:1817.
Amanat F, Stadlbauer D, Strohmeier S, Nguyen THO, Chromikova V, McMahon M, et al. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat Med. 2020;26(7):1033–6.
Voysey M, Costa Clemens SA, Madhi SA, Weckx LY, Folegatti PM, Aley PK, et al. Single-dose administration and the influence of the timing of the booster dose on immunogenicity and efficacy of ChAdOx1 nCoV-19 (AZD1222) vaccine: a pooled analysis of four randomised trials. Lancet. 2021;397(10277):881–91.
Abu-Raddad LJ, Chemaitelly H, Butt AA, Med VNEJ. National Study Group for COVID-19 vaccination. Effectiveness of the BNT162b2 Covid-19 vaccine against the B.1.1.7 and B.1.351. N Engl J Med. 385(2):1.
Bernal J, Andrews N, Gower C, Robertson C, bmj JS-, 2021 undefined. Effectiveness of the Pfizer-BioNTech and Oxford-AstraZeneca vaccines on covid-19 related symptoms, hospital admissions, and mortality in older adults in. BmjCom [Internet]. Available from: https://www.bmj.com/content/373/bmj.n1088.full. Accessed 13 May 2021.
Baden LR, El Sahly HM, Essink B, Kotloff K, Frey S, Novak R, et al. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N Engl J Med. 2021;384(5):403–16.
Madhi SA, Baillie V, Cutland CL, Voysey M, Koen AL, Fairlie L, et al. Efficacy of the ChAdOx1 nCoV-19 Covid-19 vaccine against the B.1.351 variant. N Engl J Med. 2021;384(20):1885–98.
Liu J, Liu Y, Xia H, Zou J, Weaver SC, Swanson KA, et al. BNT162b2-elicited neutralization of B.1.617 and other SARS-CoV-2 variants. Nature. 2021;596(7871):273–5.
Ramanathan K, Antognini D, Combes A, Paden M, Zakhary B, Ogino M, et al. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: an interim analysis of a randomised controlled phase 3 trial in Russia. Lancet. 2020;(January):19–21.
Gushchin VA, Dolzhikova IV, Shchetinin AM, Odintsova AS, Siniavin AE, Nikiforova MA, et al. Neutralizing activity of sera from sputnik v-vaccinated people against variants of concern (VOC: B.1.1.7, B.1.351, P.1, B.1.617.2, B.1.617.3) and Moscow endemic SARS-CoV-2 variants. Vaccines. 2021;9(7):779.
Barchuk A, Cherkashin M, Bulina A, Berezina N, Rakova T, Kuplevatskaya D, et al. Vaccine Effectiveness against Referral to Hospital and Severe Lung Injury Associated with COVID-19: A Population-based Case-control Study in St. Petersburg, Russia. medRxiv [Internet]. 2021;2021.08.18.21262065. Available from: http://medrxiv.org/content/early/2021/08/23/2021.08.18.21262065.abstract. Accessed 21 Mar 2022.
National Institute of Health. Janssen Investigational COVID-19 Vaccine: Interim Analysis of Phase 3 Clinical Data Released [Internet]. NIH News Release. 2021. Available from: https://www.nih.gov/news-events/news-releases/janssen-investigational-covid-19-vaccine-interim-analysis-phase-3-clinical-data-released. Accessed 29 Jan 2021.
Ledford H. J&J’s one-shot COVID vaccine offers hope for faster protection. Nature. 2021.
Sadoff J, Gray G, Vandebosch A, Cárdenas V, Shukarev G, Grinsztejn B, et al. Safety and efficacy of single-dose Ad26.COV2.S vaccine against Covid-19. N Engl J Med. 2021;384(23):2187–201.
Kim JH, Marks F, Clemens JD. Looking beyond COVID-19 vaccine phase 3 trials. Nat Med. 2021;27(2):205–11.
de Faria E, Guedes AR, Oliveira MS, de Godoy Moreira MV, Maia FL, dos Santos Barboza A, et al. Performance of vaccination with CoronaVac in a cohort of healthcare workers (HCW) - preliminary report. medRxiv [Internet]. 2021;2021.04.12.21255308. Available from: http://medrxiv.org/content/early/2021/04/15/2021.04.12.21255308.abstract. Accessed 15 Apr 2021.
Mallapaty S China COVID vaccine reports mixed results — what does that mean for the pandemic? Nature. 2021.
Novavax. Novavax COVID-19 Vaccine Demonstrates 89.3% Efficacy in UK Phase 3 Trial. 2021;(August 2020):3. Available from: https://ir.novavax.com/news-releases/news-release-details/novavax-covid-19-vaccine-demonstrates-893-efficacy-uk-phase-3
Callaway E, Mallapaty S. Novavax offers first evidence that COVID vaccines protect people against variants. Nature. 2021;590(7844):17.
Mahase E. Covid-19: Novavax vaccine efficacy is 86% against UK variant and 60% against South African variant. BMJ. 2021;372.
SAGE Working Group on COVID-19 vaccines. Evidence assessment: Sinopharm/BBIBP COVID-19 vaccine [Internet]. 2021. Available from: https://cdn.who.int/media/docs/default-source/immunization/sage/2021/april/2_sage29apr2021_critical-evidence_sinopharm.pdf?sfvrsn=3dfe32c1_5
Biotech B. Bharat Biotech Concludes Final Analysis for COVAXIN ® Efficacy from Phase 3 Clinical Trials. 2021;2–5.
Al Kaabi N, Zhang Y, Xia S, Yang Y, Al Qahtani MM, Abdulrazzaq N, et al. Effect of 2 inactivated SARS-CoV-2 vaccines on symptomatic COVID-19 infection in adults: a randomized clinical trial. JAMA - J Am Med Assoc. 2021;326(1):35–45.
Cohen J. COVID-19 shot protects monkeys. Science (80-). 2020;368(6490):456–7.
Gu H, Chen Q, Yang G, He L, Fan H, Deng YQ, et al. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science (80-). 2020;369(6511):1603–7.
Gretebeck LM, Subbarao K. Animal models for SARS and MERS coronaviruses. Curr Opin Virol. 2015;13:123–9.
Muñoz-Fontela C, Dowling WE, Funnell SGP, Gsell PS, Riveros-Balta AX, Albrecht RA, et al. Animal models for COVID-19. Nature. 2020;586(7830):509–15.
Sheikh A, McMenamin J, Taylor B, Robertson C. SARS-CoV-2 Delta VOC in Scotland: demographics, risk of hospital admission, and vaccine effectiveness. Lancet. 2021;397(10293):2461–2.
Riemersma KK, Grogan BE, Kita-Yarbro A, Halfmann P, Kocharian A, Florek KR, et al. Shedding of Infectious SARS-CoV-2 Despite Vaccination when the Delta Variant is Prevalent - Wisconsin, July 2021. medRxiv [Internet]. 2021;2021.07.31.21261387. Available from: https://doi.org/10.1101/2021.07.31.21261387.
Singh K, Mehta S. The clinical development process for a novel preventive vaccine: an overview. J Postgrad Med. 2016;62(1):4–11.
Wagner R, Hildt E, Grabski E, Sun Y, Meyer H, Lommel A, et al. Accelerated development of covid-19 vaccines: technology platforms, benefits, and associated risks. Vaccines. 2021;9(7).
Krause PR, Fleming TR, Longini IM, Peto R, Briand S, Heymann DL, et al. SARS-CoV-2 variants and vaccines. N Engl J Med. 2021;385(2):179–86.
Bartsch SM, O’Shea KJ, Ferguson MC, Bottazzi ME, Wedlock PT, Strych U, et al. Vaccine efficacy needed for a COVID-19 coronavirus vaccine to prevent or stop an epidemic as the sole intervention. Am J Prev Med. 2020;59(4):493–503.
Gomez PL, Robinson JM, Rogalewicz JA. Vaccine manufacturing Vaccines 6th Ed 2012;44–57.
Ball P. The lightning-fast quest for COVID vaccines - and what it means for other diseases. Nature. 2021;589(7840):16–8.
WHO. Statement for healthcare professionals: How COVID-19 vaccines are regulated for safety and effectiveness [Internet]. Who.Int. 2021. Available from: https://www.who.int/news/item/11-06-2021-statement-for-healthcare-professionals-how-covid-19-vaccines-are-regulated-for-safety-and-effectiveness
Lurie N, Saville M, Hatchett R, Halton J. Developing Covid-19 vaccines at pandemic speed. N Engl J Med. 2020;382(21):1969–73.
Acknowledgments
The authors would like to thank the Research Council at the National Institute of Genetic Engineering and Biotechnology and Tehran University of Medical Sciences for their support and provision of facilities.
Funding
This study was funded and supported by Iran University of Medical Sciences, grant number: 1399-01-97-981.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interests
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hadizadeh, N., Naderi, M., Khezri, J. et al. Appraisal of SARS-CoV-2 mutations and their impact on vaccination efficacy: an overview. J Diabetes Metab Disord 21, 1763–1783 (2022). https://doi.org/10.1007/s40200-022-01002-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s40200-022-01002-6