
Abstract
The molluscan phylum is the second specious animal group with its taxa feeding on a variety of food sources. This is enabled by the radula, a chitinous membrane with embedded teeth, one important autapomorphy. Between species, radulae can vary in their morphology, mechanical, and chemical properties. With regard to chemical composition, some taxa (Polyplacophora and Patellogastropoda) were studied extensively in the past decades, due to their specificity to incorporate high proportions of iron, calcium, and silicon. There is, however, a huge lack of knowledge about radular composition in other taxa. The work presented aims at shedding light on the chemistry by performing energy-dispersive X-ray spectroscopy analyses on 24 molluscan species, thereof two Polyplacophora, two Cephalopoda, and 20 Gastropoda, which was never done before in such a comprehensiveness. The elements and their proportions were documented for 1448 individual, mature teeth and hypotheses about potential biomineralization types were proposed. The presented work additionally comprises a detailed record on past studies about the chemical composition of molluscan teeth, which is an important basis for further investigation of the radular chemistry. The found disparity in elements detected, in their distribution and proportions highlights the diversity of evolutionary solutions, as it depicts multiple biomineralization types present within Mollusca.
Similar content being viewed by others
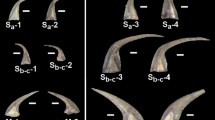
Introduction
The radula is the molluscan autapomorphy for food gathering and processing. Overall, its general shape reflects the deep molluscan phylogeny, whereas the fine morphology and chemical composition seem to be usually species-specific, serving as taxonomic character. All parameters are also, however, considered as adaptations to different ingesta types (food type, minerals, the substrate where the food is collected from, etc.) related to the ecological niche of particular species.
In most molluscan classes, the radula consists of a membrane with rows of embedded teeth, which form, together with underlain odontophoral cartilages, surrounding muscles, alary processus, and in some taxa also the jaw, the feeding apparatus, i.e. the buccal mass. The radula itself is continuously regrown by secretion of over- and underlain epithelia situated in the 'radular sac', before it is maturated in the ‘maturation zone’ and finally enters the ‘working zone’, where teeth interact with the ingesta [e.g.1,2,3].
Past studies related the radular morphology and general buccal mass anatomy with the ingesta preferred [e.g.4,5,6,7,8,9,10,11,12,13], supplemented by phenotypic plasticity studies addressing the morphological answers to shifts in the diet [e.g.14,15,16,17,18]. However, the radular tooth shape is not always a good indicator for trophic specializations19, since morphology seems to be more phylogenetically fixated and rather less adaptive in some cases12. Additionally, experimental approaches revealed that not all teeth necessarily interact with the ingesta, but still contribute to the overall radular function by e.g. reinforcing the spanned radula20,21, which indicates that tooth function and the identification of potential trophic adaptations is more complex than it has been thought before.
The mechanical properties (e.g. stiffness and hardness) of the radular teeth material can be considered as adaptive to certain ingesta types. This is well investigated for the dominant lateral teeth of Polyplacophora and patelliform gastropods, which are of high stiffness and hardness as adaptations to feeding on algae covering rocks22,23,24,25,26,27,28. For these taxa, the mechanical parameters have their origin in the architecture of the organic components (matrix of alpha chitin with associated proteins) as e.g. fiber orientation or density, the distinct folding or bonding conditions of the chitin, and the amount/distribution of the inorganic components (iron, silicon, and calcium) [e.g.23,25,27,28,29,30,31,32; for reviews see33,34,35,36].
This direct relationship between ingesta and tooth mechanical properties was recently also described for closely related paludomid gastropods from Lake Tanganyika; here species feed either on algae covering sand, mud, rock, or multiple surfaces12,37,38,39,40. The origins of the differences in hardness and stiffness remained, however, enigmatic, as radular teeth are completely understudied with regard to their structural organization and chemical composition outside the polyplacophoran or limpet realm. Only very few past studies focused on the radular composition in non-patelliform gastropods [e.g.41,42,43], cephalopods43, or scaphopods43,44,45.
We thus aim here on shedding some light on the chemical composition of radular teeth. We performed elemental analyses using energy disperse x-ray spectroscopy (EDX, EDS), overall on 1448 mature teeth of 24 species, thereof two Polyplacophora, two Cephalopoda, and 20 Gastropoda species. For the gastropods, we have chosen representative taxa from the Vetigastropoda, Neritimorpha, Heterobranchia, Caenogastropoda, and one Patellogastropoda for comparison. For the Caenogastropoda, we included six paludomid species from Lake Tanganyika, which were previously studied extensively with regard to the radular tooth mechanical properties, because one goal was the determination of the origin of heterogeneities. The elements incorporated in the mature teeth were identified, documented, and compared between the species studied. The aim of this project is the identification of similarities and differences in tooth mineralization between the molluscan taxa, which was never done before for such an elevated quantity of taxa. The results are compared and interpreted against the background of previous studies on the chemical composition of radular teeth and functional adaptations of different radulae.
Results
Radular morphology and types
Overall, the here analysed radulae are highly distinct in morphology, size, and quantity of tooth rows (see Table 1 and Supplementary Figs. 1–24 for each species). However, these differences are rather interspecific, as the selected specimens, which were of similar body size for each species, show only little intraspecific variability in the radular parameters studied here.
Based on the arrangement and quantity of certain tooth shapes, the radulae of the studied species could be assigned to the following published radular type categories (see Table 1): those of the Polyplacophora and Patellogastropoda to the docoglossan, those of the Cephalopoda to the homodont, those of the Vetigastropoda and Neritimorpha to the rhipidoglossan (special type ‘neritinomorph’ for Vittina turrita), those of the Caenogastropoda to the taenioglossan or stenoglossan, and those of the Heterobranchia Doris pseudoargus and Cornu aspersum to the isodont type. Radulae of Onchidoris bilamellata, Aeolidia papillosa, and Polycera papillosa could not be assigned to existing categories due to their diverging morphologies. The categories usually reflect phylogeny, except for the docoglossan type, which was detected in Polyplacophora and Gastropoda (Patellogastropoda). Within Heterobranchia, specifically Nudibranchia (O. bilamellata, A. papillosa, P. quadrilineata, D. pseudoargus), we detect a rather large diversity of radular shapes and thus different types. In addition, radular formulae, based on the quantity and arrangement of teeth with a specific shape, were determined. These do not consistently reflect phylogeny, except for Polyplacophora, Cephalopoda, and each of the two large Caenogastropoda groups (Buccinoidea and Paludomidae), since selected radulae are rather diverging in their interspecific morphology.
Histioteuthis spec. possesses the shortest radula (< 1 mm), Patella vulgata the longest one (37 mm). Faunus ater, Haliotis tuberculata, V. turrita, and Littorina littorea show radulae that are of 10–23 mm length, all other species of 1.4–10 mm. Areas of radulae in Buccinum undatum, V. turrita, C. aspersum, and P. vulgata are rather large (14–35 mm2). The largest one was detected for H. tuberculata (59 mm2) and the smallest for H. spec. (0.39 mm2). Most species have less than 100 tooth rows, with A. papillosa possessing the smallest quantity (9 rows). The radulae of Rochia conus, Lavigeria nassa, H. tuberculata, Spekia zonata, F. ater, C. aspersum, Reymondia horei, and P. vulgata contain 100–200 rows. V. turrita’s radula possesses 149–151 rows, whereas L. littorea’s one has the highest number of rows (280–281). Length, area, and tooth row number do not seem to relate to the phylogeny as closer related species (e.g. the Paludomidae: L. nassa, L. grandis, R. horei, S. zonata, and C. johstoni) show large differences in these parameters.
Ingesta
Overall, the preferred ingesta types (see Table 1) vary greatly within classes, especially within Vetigastropoda, Caenogastropoda, and Heterobranchia, and convergent adaptations to similar trophic categories are present. The two studied Polyplacophora species (Acanthochitona fascicularis and Lepidochitona cinerea), the Patellogastropoda Patella vulgata, the Vetigastropoda Rochia conus, the Caenogastropoda Lavigeria grandis, L. nassa, Reymondia horei, and Spekia zonata can be regarded as solid ingesta feeders. Neritimorpha Vittina turrita and Caenogastropoda Littorina littorea seem to forage on solid, but also medium hardness ingesta, the Vetigastropoda Haliotis tuberculata and the Heterobranchia Onchidoris bilamellata, Polycera quadrilineata, and Doris pseudoargus on ingesta of medium stiffness and hardness. The Caenogastropoda Cleopatra johnstoni and the Heterobranchia Aeolidia papillosa were the only species foraging on softer ingesta. The Cephalopoda Histioteuthis spec. and Loligo vulgaris, the Caenogastropoda Paramelania damoni, Faunus ater, Anentome helena, and Buccinum undatum, as well as the Heterobranchia Cornu aspersum seem to forage on the widest range of food types (soft to solid).
Elements detected
Overall, the presence of many elements is rather puzzling and does not follow, in each case, the phylogeny (see Figs. 1 and 2; Supplementary Figs. 1–27; Supplementary Table 3 for mean, SD, sum of means, and N): S was detected in the radula of each studied species. Mg, Ca, Na, and P in most, but in Cornu aspersum no Mg, Na, and P and in Cleopatra johnstoni no Ca was found. Cl was determined for radulae of Acanthochitona fascicularis, both Cephalopoda, Vittina turrita, all Caenogastropoda, Onchidoris bilamellata, and Aeolidia papillosa. F was detected in A. fascicularis, Patella vulgata, both Vetigastropoda, Littorina littorea, O. bilamellata, and Doris pseudoargus. Si was found in Lepidochitona cinerea, both Cephalopoda, P. vulgata, both Vetigastropoda, V. turrita, all Caenogastropoda except C. johnstoni and Buccinum undatum, and in Cornu aspersum. Cu was determined for radulae of both Cephalopod species and Reymondia horei. K was found in both Polyplacophora, P. vulgata, Paramelania damoni, C. johnstoni, R. horei, Paludomus siamensis, L. littorea, and O. bilamellata.

Elemental proportions of the species’ radular teeth, summarized for taxa (Patellogastropoda, Polyplacophora, Vetigastropoda, Caenogastropoda, Cephalopoda, Heterobranchia, Neritimorpha) to ease comparison in the radular mineral content.

Elements detected, the mean atomic percent (represented by different colours), and assigned ingesta category (soft, medium, solid, soft to solid, medium to solid) for each species, sorted to the major molluscan groups (Polyplacophora, Cephalopoda, Gastropoda: Patellogastropoda, Vetigastropoda, Caenogastropoda, Neritimorpha, Heterobranchia). Elements are sorted to the defined composition-types (I, II, III, IV, V, OB).
Elemental proportions
Summarizing all radulae studied (for mean, SD, and N see Supplementary Table 1), we detect that Fe is present in the highest proportions, followed by Si, Ca, P, F, Na, Mg, S, Cl, Cu, and finally K. Of the 1448 areas studied, most of them contained Ca, followed by P, Mg, S, Cl, Na, Si, F, Fe, Cu, and finally K.
Overall, the highest content of all analysed elements (sum, in atomic %, of the means of F, Na, Mg, Si, P, S, Cl, K, Ca, Fe, and Cu; see Supplementary Table 2 for values) was detected for the studied Patellogastropoda. This is followed by the Polyplacophora, Heterobranchia, Caenogastropoda, Cephalopoda, Vetigastropoda, and finally the Neritimorpha.
Composition- and biomineralization-types
Overall, we detect that multiple composition- and biomineralization types are present within each individual species and also within individual teeth (see Fig. 2). Within Polyplacophora and Patellogastropoda we detected strong indications for the presence of the composition type I (containing Fe), II (Mg, Ca), III (Ca, P, Cl, F), IV (Si), OB (Na, S, K). In Cephalopoda, the types II, III, IV, V (Cu), and OB occur. In Vetigastropoda and Neritimorpha, the types II, III, IV, and OB were detected, in Vittina turrita—additionally the type I. In Caenogastropoda and Heterobranchia, the radular composition greatly varies between taxa. Overall, in Caenogastropoda, all composition types were found. However, in each species, the types II, III, and OB were always present, whereas Fe was only determined in Reymondia horei and Littorina littorea, Cu — in R. horei, and K — in Paramelania damoni, Cleopatra johnstoni, R. horei, Faunus ater, and L. littorea. C. johnstoni is the only species that seems to lack Ca. In Heterobranchia, the types II and OB were found in each species. The type III (apatite) is present in Onchidoris bilamellata, Aeolidia papillosa, Polycera quadrilineata, and Doris pseudoargus, but not in Cornu aspersum, as P, Cl, and F were not determined in this species. Si was only detected in C. aspersum and K in O. bilamellata.
Ingesta versus radular morphology and elemental proportions
Morphology
The longest radulae were detected in species foraging on medium to solid ingesta, followed by solid-, medium-, soft-to-solid-, and finally soft-feeders with the shortest radulae (see Table 2 and Supplementary Figs. 28–30). The largest radular area was calculated for medium-, medium-to-solid-, soft-to-solid-, solid-, and finally for the soft-feeders with the smallest area. Species foraging on medium-to-solid ingesta possess the highest quantity of tooth rows, followed by solid-, soft-to-solid-, medium-, and finally with the least quantity of tooth rows the soft-feeders.
All elements
In general, we detect that radulae of species foraging on solid ingesta possess the highest proportions of all studied elements, followed by species foraging on medium, medium-to-solid, soft-to-solid, and finally species feeding on soft ingesta.
Composition-type I
The highest Fe-proportions (means) were detected in the exclusively solid-, followed by the medium-to-solid-feeders. No Fe was detected for all other ingesta types.
Composition-type II and III
The highest proportions of Mg were detected in species foraging on medium ingesta, followed by solid-, soft-, medium-to-solid-, and finally soft-to-solid-feeders. Ca was detected in the highest proportions in the medium-, followed by the soft-, solid-, soft-to-solid-, and medium-to-solid-feeders. P was mainly found in species feeding on medium ingesta, followed by soft-to-solid-, solid-, medium-to-solid-, and finally soft-feeders. Cl was detected in the highest proportions in medium-feeders, followed by species feeding on soft-to-solid, soft, solid, and finally medium-to-solid ingesta. The highest proportions of F were found in radulae of medium-, followed by solid-, and finally medium-to-solid-feeders. In soft and soft-to-solid-feeders, no F was found.
Composition-type IV
The highest Si-content was detected in solid-, followed by medium-, medium-to-solid-, and finally soft-to-solid-feeders. No Si was found in species feeding on soft ingesta.
Composition-type V
The highest Cu proportions were detected in the solid-feeder Reymondia horei and less in the cephalopods foraging on soft-to-solid ingesta. All other radulae seem to lack Cu.
OB
S was detected in the highest proportions in species feeding on soft, followed by solid, soft-to-solid, medium-to-solid, and finally medium ingesta. The highest Na-proportions were detected in solid-, followed by the soft-, soft-to-solid-, medium-, and finally medium-to-solid-feeders. K was detected in the highest proportions in solid-, followed by medium-, medium-to-solid-, soft-to-solid-, and finally soft-feeders.
Correlations between parameters
In some cases we could detect correlations (please see Supplementary Tables 4–16 for correlation coefficients). In general, radular area highly correlates with radular length and radular width. Additionally, in most cases, proportions of all elements highly correlate with each individual element.
For individual elemental proportions, we here only highlight some, but the picture is rather puzzling. When all species are pooled together (see Supplementary Table 4) Ca and Cl, F and Ca, Fe and Ca, P and Ca, P and Cl, Si and Ca, Si and Fe correlate highly. For all soft ingesta feeders pooled together (see Supplementary Table 5) Mg and Na, K and P, Ca and P, Cl and P, Cl and K, Cl and Ca highly correlate. For all soft-to-solid ingesta feeders pooled together (see Supplementary Table 6) K and P, K and S, Ca and K, Cl and P, Cl and K highly correlate. For all medium ingesta feeders pooled together (see Supplementary Table 7) S and Si, K and Mg, Ca and P, F and P, F and Ca, Cl and S highly correlate. For all medium-to-solid ingesta feeders pooled together (see Supplementary Table 8) Si and Na, Si and Mg, K and Na, Ca and P, Ca and K, F and P, F and K, F and Ca, Cl and P, Cl and Ca highly correlate. For all solid ingesta feeders pooled together (see Supplementary Table 9) Si and Mg, P and Si, S and Si, K and P, Ca and P, Cl and P correlate highly.
When the species studied are sorted by their taxonomic group we find, for most parameters no high correlations. However, for Caenogastropoda (see Supplementary Table 10) K and Si, F and P, F and K, F and Ca, Cl and P highly correlate. In Cephalopoda (see Supplementary Table 11) Cl and Ca correlate highly. In Heterobranchia (see Supplementary Table 12) K and Mg, Ca and P, F and P, Cl and Na are highly correlated. In Neritimorpha (see Supplementary Table 13) Si and Na, Si and Mg, Ca and P, Cl and P, Cl and Ca are highly correlated. In Patellogastropoda (see Supplementary Table 14) Si and Mg, P and Mg, S and P, K and Na, K and P, K and S, Ca and Mg, Ca and P highly correlate. In Polyplacophora (see Supplementary Table 15) P and Na, S and Mg, K and P, Fe and Si, F and K, Cl and Na are highly correlated. In Vetigastropoda (see Supplementary Table 16) S and Na highly correlate.
PCA on the individual parameters (elemental proportions, radular length, radular width, radular area, tooth rows) for all species pooled together detected no clustering (see Supplementary Fig. 31 A with highlighted systematic groups and B with highlighted ingesta categories).
Discussion
A detailed list of previous studies aimed at determining the chemical composition of the molluscan radula is provided in Supplementary Table 17. Most of the previous research has been done on the Polyplacophora with the focus exclusively on the dominant lateral teeth [for reviews see33,34,35,36,80], except for one study on Lepidochitona cinerea determining the elemental composition of all tooth types32. Many of these analyses focused on the Fe biomineralization and the phase transformations during maturation [e.g.23,29,81,82,83,84,85,86,87,88,89]. Overall, in previous studies F, Na, Mg, Si, P, S, Cl, K, Ca, Fe, and Cu was detected in the dominant lateral teeth (= lateral teeth II) of Polyplacophora. For Lepidochitona cinerea, in our previous paper, we did not detect Cl, F, and Cu and in Acanthochitona fascicularis — no Si and Cu.
The following Fe proportions of mature dominant lateral teeth were previously determined in Polyplacophora: for A. fascicularis — 59.2% 90, 62%85,91 or few percent92 were detected. For Plaxiphora — 86.6%90, 17–27%85, or 26.7%93; for Cryptochiton — 51.8%81 or 69%29; for Ischnochiton — 62%85; for Onithochiton — 66%85 or 0.2%93; for Cryptoplax — ~ 90 weight % in the cap, ~ 30 weight % in the core, junction zone, and basis83; and for Chiton — 97%94 were detected. For mature L. cinerea, we previously32 found Fe proportions of maximal 32% (atomic ratio, atomic %) and for A. fascicularis — maximal 29% in the dominant lateral teeth.
For Polyplacophora, P was previously reported81,83,85,93,95 in form of iron phosphate86,96,97 or apatitic calcium phosphate92,93,95,98,99,100. F related to Ca100,101 was also previously reported for the dominant lateral teeth of chitons. In Acanthopleura85,92 and Onithochiton85, Ca was abundant to maximal ~ 30 atomic % and P to maximal ~ 20 atomic %. For Lepidochitona cinerea, we detected Ca in proportions of maximal 8% and P to 7% and for Acanthochitona fascicularis to maximal 6% and P to 9%.
Additionally, Si83,85,98,99 and Mg85,93 were previously detected in the dominant lateral teeth of chitons. S was also previously determined83. It is associated with the tanning of the organic matrix and with the appearance of proteins82. Additionally,102 detected Zi, K, F, S, Na, and Cl in radular segments of Clavarizona90, Ca, P, Mg, S, Na, Zi, K, Al, Cu, and Si in radulae of Acanthopleura and Plaxiphora, and83 Mg (with max ~ 5.5 weight %), K (with max ~ 1.0 weight %), Na (with max ~ 2.0 weight %), Si (with max ~ 1.0 weight %), Al (with max ~ 0.5 weight %), and S (with max ~ 0.8 weight %) in Cryptoplax. These elements, except for Zi and Cu, which were not found, occurred in smaller proportions (0–5%) in both Lepidochitona cinerea and Acanthochitona fascicularis. For the central, lateral I, and marginal teeth we detected less minerals than in the dominant lateral teeth in both species.
In Cephalopoda, only one study on the radular chemistry exists, to the best of our knowledge. In Octopoda43 targeted, but did not detect Si and Fe. We here determined Na, S, Cu, Si, P, Cl, Ca, and Mg in the radula of Histioteuthis spec. and Loligo vulgaris with proportions < 4%.
Within the Gastropoda, the Patellogastropoda received the most attention41,42,43,90,103,104,105,106,107,108,109,110,111,112,113,114,115. However, few studies focused on the overall radular composition, since most analyses, e.g. ashing and treatments with different acids or Raman spectroscopies, EDX, rather targeted the presence and crystalline shape of Fe and Si [e.g.41,103,104,106,107,108,109,110,111,114]. In fewer studies, other elements were of interest [e.g.42,43,90,105,112,113]. Overall, in Patellogastropoda, Na, Mg, Si, P, Cl, K, Ca, Fe, Cu, and S were previously found. For Patella vulgata, we here detected Na, Mg, Si, P, K, Ca, Fe, S, but no Cu and Cl. We additionally determined F. Similar to previous studies, we detected Si and Fe in high proportions in the dominant teeth (18–38%), whereas all other elements were present in smaller proportions only.
For the remaining gastropod taxa, only few analyses on the radular chemistry were conducted and usually the presence of elements, but not their proportions, could be determined. One of the earliest studies was done by41 depicting results from Bergh, who performed complex chemical analyses of ashing and dissolving radulae from the Caenogastropods Charonia lampas (detecting P, Ca, and Fe), Lamellaria perspicua (detecting no Si), and Gibberulus gibberulus (probably detecting none of these elements, this is not clear) in different acids. Additionally,41 presented his own results on the radulae of the Caenogastropod Tonna galea and the Heterobranch Helix nemoralis discovering P, Ca, and Fe in both by employing the same experiment. Sollas42 was the first, who studied the radular chemistry in an elevated quantity of taxa, and43 proceeded. Overall, their protocols are rather complex, involving analytical chemistry methods (ashing, staining, boiling, treating with acids, and using diffusion column) or physics (radula’s refractive index). Sollas42 determined rather the presence of elements and43 specifically tested the occurrence of Si and Fe. For the Caenogastropoda Potamopyrgus antipodarum, Lacuna vincta, Murex branchialis, and Aporrhais pespelecani, the Heterobranchia Scaphander lignarius, Aplysia punctata, and Jorunna tomentosa,43 determined no Si and no Fe. For the Heterobranchia (Cornu aspersum),42 determined Si and P. She detected Si in specimens collected during winter and phosphoric acid (P) in specimens collected during spring.116 performed EDX analyses on five specimen of C. aspersum detecting Ca in all specimen and Si in one, even though specimens were also inventoried in spring depicting the inconsistency of elements embedded. For the Vetigastropoda (Haliotis tuberculata),42 detected Si, Ca, and Fe.
The following species were studied by42 and43, but for many species their results are contradictory. In the Caenogastropoda Littorina littorea42 detected Mg, P, Ca, and Fe, whereas43 found no Ca and no Fe. Nucella lapillus and Buccinum undatum were also studied by42, but the results are not clear from the publication, and43 detected no Si and no Fe in both species. For the Vetigastropoda (Emarginula fissure and Calliostoma zizyphinum)42 detected P, Ca, and Fe, whereas43 determined the absence of Fe and Si. Then117 and26,118 were the first to close the existing gap in knowledge about the radular composition of Vetigastropoda. Gray117 detected Na, Mg, Si, Cl, Ca, and Fe (EDX), and26,118 Mg, Cl, Ca, and Fe (by EDX and inductively coupled plasma-optical emission spectrometry) in the limpet Megathura crenulata. Within Vetigastropoda, we detected Na, S, Cu, Si, P, Cl, Ca, and Mg; all of them in low proportions < 2%. Cu and S were not documented before, whereas Fe was detected in previous studies117,118. For the Neritimorpha, only one past study addresses the mineral content detecting S, Cl, K, Ca, Mg, Si, and Fe119. We additionally detected Na and P in Vittina turrita; all elements are abundant in very low proportions (< 4%). In the Caenogastropoda, we detected Fe, Mg, Ca, Cl, P, F, Si, Cu, S, Na, and K. Cu, F, Na, Si, S, and Cl were not determined before. In all species, proportions are < 6%. For the Heterobranchia, we detected more elements (Mg, Ca, Cl, P, F, Si, S, Na, K) than described in past publications41,42,43,116. Mg, Cl, F, S, Na, and K were not detected before. All elements are abundant at proportions < 15%.
Overall, the above data depicts that it is rather difficult to compare the percentages measured between studies, because in some weight percentages and in others atomic ratios were determined. Besides, methodology, sample preparation, and the analysed sample itself (whole radula or individual radular parts) differs. In addition, the presence and abundance of elements could potentially be influenced by the food available (e.g. plants containing or lacking Si) or by the chemistry of the saliva. In some taxa, specifically carnivorous gastropods, the saliva is acid [e.g.120,121], so potentially the contact of the outermost radular teeth with the saliva leads to reduced proportions. Both ideas await further research.
The generally accepted hypothesis on radular mineralization evolution states that all gastropods — besides Patellogastropoda, Neritimorpha, and Vetigastropoda — probably lack Fe in the radula [e.g.122,123,124]. However, Fe was detected previously in gastropod species [for Tonna galea, Charonia lampas, and Helix nemoralis see41, for Littorina littorea see42] and our own analyses determined it in Reymondia horei and Littorina littorea. Thus, this means that iron is not lacking, rather its proportions are reduced in these gastropod lineages (see Fig. 1).
Previous studies relate the radular length to the ingesta type. Herbivorous taxa were found to possess longer radulae than carnivorous ones125. Littorinid species, feeding on algae covering rocks, were found to possess longer radulae than species feeding from plant surface126,127,128,129,130. For Patella species, it was determined that the radular length increases with increasing usage and wear131 and, when algae are less abundant and the radula must thus be used more frequently to obtain the food necessary, its length increases132.
In general, we detected a similar pattern for the species studied here as the longest radulae with the highest quantity of tooth rows were found in species foraging on harder ingesta types (medium-to-solid, solid, medium) and the shortest ones in soft-substrate feeders. We, however, could not directly relate herbivory with longer radulae and carnivorous feeding with shorter ones. We additionally detected some relationship between radular length and proportions of elements (e.g. in Patella vulgata), so potentially more mineralized radulae are longer, because their maturation and mineralization requires more time and a longer contact to the overlain epithelia in the radular sac and mineralization zone. However, this does not seem to be the case for every species, as Lepidochitona cinerea and Acanthochitona fascicularis have relatively short heavily mineralized radulae. Thus, in these polyplacophoran species, the overlain epithelia can presumably incorporate more minerals at the same time or the radular replacement rate is faster in P. vulgata in contrast to the one in the Polyplacophora. Unfortunately, the radular replacement rate is known for few taxa: for Polyplacophorans (Acanthopleura, Plaxiphora, Patelloida, Mopalia), a rate of 0.36–0.80 rows per day was determined90,133,134 and for P. vulgata, a rate of 1.5 rows/day was described135. In Caenogastropoda, for Lacuna (Littorinidae), the rate of 3 rows/day136, for three Littorina species (Littorinidae) — 5–6 rows/day depending on the temperature2,135, and for Pomatias elegans— 5.02 rows/day2 was determined. For Heterobranchia, the rate of 2.9 rows/day in Lymnaea stagnalis 137, 5.02 rows/day in Agriolimax reticulatus2, 3.6 rows/day in adult Helix aspersa (= Cornu aspersum) was detected2. For Cepaea nemoralis, the whole radula was found to be renewed within 30–35 days138. Thus, in general, a higher degree of mineralization is inversely related to the higher replacement rate (teeth that possess larger proportions of minerals are replaced slower). However, radular replacement seems to depend on many factors, such as water temperature, metabolic rates, or age of animals135,136,139,140. Further studies on these questions are required.
In general, we detected that radulae of species, foraging on the solid ingesta, possess heavy mineralized teeth and species feeding on the soft ingesta show the smallest proportions. In biological materials, heterogeneities can have their origin in geometry, chemistry, and/or structure [for a review see141]. In the dominant lateral teeth of chitons and limpets they have their origin in the distribution of the inorganic components and in the architecture of organic components23,25,27,28,29,30,31. We have previously correlated the hardness and elasticity values in Lepidochitona cinerea with the iron and the calcium proportions32, which was previously also described for limpet teeth24,104,108,142 and for other chitons23,25,29,30. For the paludomid gastropods, we previously measured elasticity modulus values ranging from 2 GPa at the tooth basis to 8 GPa in the cusp in solid substrate feeders, whereas soft substrate feeders possessed significantly softer teeth (4.6 GPa)37,38,40. In these species, we here detected inorganic elements in rather small proportions. We thus propose that specific cross-linking conditions of the chitin due to tanning1, fiber arrangement, and density22,23,26,28,31,88,143,144,145 rather cause the heterogeneities in mechanical properties. We previously also detected that the capability of wet teeth to rely on one another and to redistribute the mechanical stress increases the radula’s resistance to structural failure in paludomid gastropods146,147. This altogether probably enables the feeding on harder ingesta types. Whether these mechanisms are also applicable for the other molluscan species, await further investigations.
Methods
Specimen studied and dissection
Mollusks were obtained from various sources (see Table 1 for details): individuals of Littorina littorea and Lepidochitona cinerea were collected at the North Sea in summer 2019 and those of Patella vulgata in autumn 2020. The gastropods Cornu aspersum, Rochia conus, Haliotis tuberculata, Vittina turrita, Faunus ater, and Anentome helena were bought from online pet shops. All of them were shortly boiled and preserved in 70% ethanol. Individuals of Aeolidia papillosa, Onchidoris bilamellata, Polycera quadrilineata, and Doris pseudoargus were received from the Biologische Anstalt Helgoland in May 2021, kept in aquarium for 2 weeks in Hamburg, before the gastropods naturally died and then were preserved in 70% ethanol. Frozen specimens of Loligo vulgaris were bought from Fische Schmidt (store specialized on edible fish, Eppendorfer Baum 18, 20,249 Hamburg) for the dissection course of the Universität Hamburg, radulae were directly extracted from defrozen squids and preserved in 70% ethanol. Samples of Lavigeria grandis, L. nassa, Paramelania damoni, Cleopatra johnstoni, Reymondia horei, Spekia zonata, Paludomus siamensis, Buccinum undatum, Acanthochitona fascicularis, Histioteuthis sp. were extracted from already preserved (70% ethanol) specimens, some of them already inventoried in museum collections (Zoologisches Museum Hamburg, ZMH; Museum für Naturkunde Berlin, ZMB).
Species identification was reviewed by employing the relevant literature, the nomenclature and systematic position were checked using molluscabase.org. Not previously inventoried specimens were incorporated in the malacological collection of the ZMH, which is now part of the Leibniz-Institut für die Analyse des Biodiversitätswandels (LIB).
Overall, data from 72 adult specimens were analysed for this study. For each species, three adult specimens were selected, except for Histioteuthis spec. with two. We have chosen specimens of similar size per species, since the relationship between specimens’ length and radular size is puzzling. Some previous studies relate both parameters148 and others rather see a loose relationship or could not relate them for every species125,149. Additionally, the specimens chosen for each species were collected at the same time since seasonal dependencies in radular length were previously reported18. All data presented here is new, except for the elemental composition and radular morphology of Lepidochitona cinerea, which was taken from32. In this previous study, we analysed the ontogeny of the elemental composition and the mechanical parameters hardness and elasticity in three specimens of L. cinerea. For the present study, we included only the data from the working zone (the mature part) for the purpose of comparison between species.
Habitus images were either taken employing the Keyence Digital Microscope VHX-5000 (KEYENCE, Neu-Isenburg, Germany) or by using an iPad Pro (11 zoll; Apple Inc., Cupertino, USA) equipped with a 12-megapixel wide angle lens. Each specimen was dissected, the radula was carefully extracted by tweezers and then manually freed from surrounding tissue.
Scanning electron microscopy (SEM)
For images of the whole radula or the radular working zone, radulae (three per species, except for Histioteuthis spec. with two) were cleaned in an ultrasonic bath for 2–20 s and afterwards arranged on scanning electron microscopy (SEM) sample holders (see Supplementary Figs. 1–24). All radulae were first documented with the Keyence Digital Microscope VHX-5000 or VHX-7000 (KEYENCE, Neu-Isenburg, Germany). Here the length and width of each radula were measured and the quantity of tooth rows counted. From the length and width, the radular area was calculated. Two radulae per species were then visualized uncoated employing the Tabletop Microscope TM 4000 Plus (Hitachi, Tokyo, Japan) for more detailed images and one radula per species was coated and documented with the Zeiss LEO 1525 (One Zeiss Drive, Thornwood, USA) to receive images with a very high resolution (except for images of L. cinerea, they were taken from150, and of H. spec., as their radulae were documented uncoated and afterwards used for the EDX). Based on the morphology and arrangement of teeth, which were also categorized (e.g. central tooth, lateral tooth I, lateral tooth II, marginal tooth, inner teeth, outer teeth, etc.) according to their shape, size, and position on the membrane, radulae were assigned to different radular types (e.g. docogloss, isodont, rhipidogloss, etc.), if a suitable category could be determined from literature [e.g.5,151,152,153,154]. Then, the radulae, which were previously documented uncoated (two per species), were rewetted with 70% ethanol and loosened from the SEM sample holder and used for elemental analysis.
Elemental analysis (EDX)
Wet radulae were arranged on glass object slides (Carl Roth, Karlsruhe, Germany) with double-sided adhesive tape. They were positioned along their longitudinal axis so that the outermost teeth of one side were directly attached to the slide. The adjacent and more inner teeth were located above, followed by the central teeth, the inner teeth from the other side, and finally, on top, outer teeth again. Each radula was then dried for three days under ambient temperature and afterwards surrounded with a small, metallic ring ensuring an almost parallel sample surface. Epoxy resin (RECKLI EPOXI WST, RECKLI GmbH, Herne, Germany) was filled into the metallic ring and left polymerizing at room temperature for three days. This specific epoxy was chosen, since it does not infiltrate the teeth. Object slide and tape were then removed and, to receive longitudinal sections of each tooth, the embedded radulae were polished until the outer teeth were on display (controlled by examining the samples in the light microscope) using sandpapers of different roughness. Then samples were smoothed with aluminium oxide polishing powder suspension of 0.3 μm grainsize (PRESI GmbH, Hagen, Germany) on a polishing machine (Minitech 233/333, PRESI GmbH, Hagen, Germany). After polishing, the samples were cleaned from the polishing powder by an ultrasonic bath lasting five minutes. Samples were then coated with platinum (5 nm layer) and the elemental compositions of specific areas of the embedded teeth were examined employing the SEM Zeiss LEO 1525 (One Zeiss Drive, Thornwood, New York, USA) equipped with an Octane Silicon Drift Detector (SDD) (micro analyses system TEAM, EDAX Inc., New Jersey, USA) always using an acceleration voltage of 20 keV and the same settings (e.g. lens opening, working distance, etc.). Before measuring a sample the detector was always calibrated using cupper. We performed elemental mappings for test purposes, but for elements that are present in rather lower proportions, this method is not sensitive enough. We thus focused on the elemental analysis of small areas (10–400 μm2, depending on the tooth) trying to analyse the largest possible area.
The elements H (hydrogen), C (carbon), N (nitrogen), O (oxygen), Pt (platinum), Al (aluminium), Ca (calcium), Na (sodium), Mg (magnesium), Si (silicon), P (phosphorus), S (sulphur), Cl (chlorine), K (potassium), F (fluorine), Cu (copper), and Fe (iron) were detected and their proportions measured. We used the data of atomic ratio (atomic %) for this study. These values were received with two positions after the decimal point, lower proportions were not detectable with this method and therefore they were given as 0.00. We did not analyse and discuss the following elements, as they are either the elemental basis of chitin (H, C, N, O), the coating (Pt), or the polishing powder (Al, O).
After analysing the outer teeth, each sample was again polished and smoothened until the next tooth type was on display; cleaning procedure and EDX analyses were again performed. These steps were repeated until all teeth were analysed. In this study, we present the results of the radular working zone, which is not covered by epithelia. Thus, all teeth are mature. Overall, we use data of 1448 analysed teeth from 49 specimens.
Statistical analyses
All statistical analyses (mean, standard deviations) and visualizations with boxplots, pie charts, or trend lines were performed with JMP Pro, Version 14 (SAS Institute Inc., Cary, NC, 1989–2007). Correlation coefficients and PCA were also conducted in JMP.
Composition- and biomineralization-types
With EDX analysis, the proportions of the individual elements, present in a defined area, can be documented, whereas the specific bonding and structure of molecules cannot be analysed. However, from the percentile occurrence, in comparison with past studies on the radular chemical composition involving, we propose that the elements, detected here, are potentially part of the following molecules or minerals. These were assigned to different composition- or biomineralization-types:
Category Type 1
Characterized by the presence of Fe. Potentially present in the form of magnetite as documented in polyplacophoran or goethite in limpets [e.g.30,31,99,109,155,156,157,158,159].
Category Type 2
Characterized by the presence of Mg and Ca. Elements are potentially involved in the protein packing, an increase in density of chitin fibres and in material stiffness as documented in limpet teeth26.
Category Type 3
Characterized by the presence of Ca, P, Cl and/or F. These elements (Ca:P:Cl/F) occur approximately in these ratios to one another: 5:3:1. They are potentially part of apatite, either fluorapatite, Ca5[F|(PO4)3], or chlorapatite, Ca5[Cl|(PO4)3] as previously described for radular teeth of polyplacophorans [e.g.85,90,91,95,98,160,161,162].
Category Type 4
Characterized by the presence of Si. Potentially present in the form of silica as documented in limpet teeth [e.g.80,115].
Category Type 5
Characterized by the presence of Cu.
Category OB (organic bonds)
The presence of Na, K, and S is often related to the protein bonding [e.g.163,164].
Ingesta categories
The precise trophic preference of molluscan species is completely understudied. Some past approaches on various molluscan species determining the food preferences were based on the analysis of ingesta consumption, grazing activity, food choice experiments, stomach content, and faecal analyses [e.g.10,165,166,167,168,169,170,171,172,173,174,175,176,177]. These studies overall show, that very various food types can be intaken by one individual species. However, this cannot be generalized, since some taxa are less flexible with regard to the ingesta intaken [e.g. Nudibranchia, see178].
For this study, we defined ingesta categories that are rather broad, because (a) either the species’ food preference has only been described anecdotally and (b) even if the specific food type was known, its precise mechanical properties and its structural resistance to feeding is difficult to determine [see179,180] and that is why these properties remain unknown. We collated the data on ingesta from the literature, if available, and assigned it to the following ingesta categories (please, see Table 1 for food types per species and corresponding references):
Soft Algae from sand or mud, sea anemones.
Medium Macro algae, soft parts of barnacles, encrusted Bryozoa, or Porifera.
Solid Algae/plants from rocks and/or cora ls.
Soft to solid Fish, crustaceans, and cephalopods; algae from rocks and sand; gastropods, fish eggs, shrimps, and carrion; Polychaeta, fish eggs, bivalves, carrion, etc.
Medium to solid Algae, fleshy macro algae, also from rocks.
Systematics
The cladogram, depicting the systematic position of the species studied, was created by combining data from published phylogenies12,181,182,183,184,185,186,187,188,189 with data on the latest systematic position of the species from molluscabase.org.
Human or animals rights
For this article no research was conducted on live vertebrates and/or higher invertebrates.
Data availability
All data is provided in the supplementary material.
References
Runham, N. A study of the replacement mechanism of the pulmonate radula. J. Cell Sci. 3(66), 271–277 (1963).
Runham, N. & Isarankura, K. Studies on radula replacement. Malacologia 5, 73 (1966).
Mackenstedt, U. & Märkel, K. Radular structure and function. In The Biology of Terrestrial Molluscs (ed. Barker, G. M.) 213–236 (CABI Publishing, Oxon, United Kingdom, 2001).
Crampton, D. M. Functional anatomy of the buccal apparatus of Onchidoris bilamellata (Mollusca: Opisthobranchia). Trans. Zool. Soc. Lond. 34(1), 45–86 (1977).
Steneck, R. S. & Watling, L. Feeding capabilities and limitation of herbivorous molluscs: A functional group approach. Mar. Biol. 68(3), 299–319 (1982).
Jensen, K. R. Evolution of the sacoglossa (Mollusca, Opisthobranchia) and the ecological associations with their food plants. Evol. Ecol. 11, 301–335 (1997).
Nishi, M. & Kohn, A. J. Radular teeth of Indo-Pacific molluscivorous species of Conus: A comparative analysis. J. Molluscan Stud. 65(4), 483–497 (1999).
Duda, T. F., Kohn, A. J. & Palumbi, S. R. Origins of diverse feeding ecologies within Conus, a genus of venomous marine gastropods. Biol. J. Linn. Soc. Lond. 73, 391–409 (2001).
von Rintelen, T., Wilson, A. B., Meyer, A. & Glaubrecht, M. Escalation and trophic specialization drive adaptive radiation of freshwater gastropods in ancient lakes on Sulawesi, Indonesia. Proc. R. Soc. B 271, 2541–2549 (2004).
Ekimova, I. et al. Diet-driven ecological radiation and allopatric speciation result in high species diversity in a temperate-cold water marine genus Dendronotus (Gastropoda: Nudibranchia). Mol. Phylogenet. Evol. 141, 106609 (2019).
Mikhlina, A., Ekimova, I. & Vortsepneva, E. Functional morphology and post-larval development of the buccal complex in Eubranchus rupium (Nudibranchia: Aeolidia: Fionidae). Zoology 143, 125850 (2020).
Krings, W. Trophic specialization of paludomid gastropods from 'ancient' Lake Tanganyika reflected by radular tooth morphologies and material properties, Thesis, Universität Hamburg (2020).
Krings, W., Brütt, J.-O., Gorb, S. N. & Glaubrecht, M. Tightening it up: Diversity of the chitin anchorage of radular-teeth in paludomid freshwater-gastropods. Malacologia 63(1), 77–94 (2020).
Bleakney, J. S. Indirect evidence of a morphological response in the radula of Placida dendritica (Alder & Hancock, 1843) (Opisthobranchia: Ascoglossa/ Sacoglossa) to different algae prey. Veliger 33(1), 111–115 (1990).
Jensen, K. R. Morphological adaptations and plasticity of radular teeth of the Sacoglossa (= Ascoglossa) (Mollusca: Opisthobranchia) in relation to their food plants. Biol. J. Linn. Soc. Lond. 48, 135–155 (1993).
Reid, D. G. & Mak, Y.-M. Indirect evidence for ecophenotypic plasticity in radular dentition of Littorina species (Gastropoda: Littorinidae). J. Molluscan Stud. 65, 355–370 (1999).
Padilla, D. K., Dilger, E. K. & Dittmann, D. E. Phenotypic plasticity of feeding structures in species of Littorina. Am. Zool. 40, 1161 (2000).
Ito, A., Ilano, A. S., Goshima, S. & Nakao, S. Seasonal and tidal height variations in body weight and radular length in Nodilittorina radiata (Eydoux and Souleyet, 1852). J. Molluscan Stud. 68, 197–203 (2002).
Padilla, D. K. Form and function of radular teeth of herbivorous molluscs: Focus on the future. Am. Malacol. Bull. 18(1/2), 163–168 (2003).
Krings, W. & Gorb, S. N. Substrate roughness induced wear pattern in gastropod radulae. Biotribology 26, 100164 (2021).
Krings, W., Hempel, C., Siemers, L., Neiber, M. T. & Gorb, S. N. Feeding experiments on Vittina turrita (Mollusca, Gastropoda, Neritidae) reveal tooth contact areas and bent radular shape during foraging. Sci. Rep. 11, 9556 (2021).
Lu, D. & Barber, A. H. Optimized nanoscale composite behaviour in limpet teeth. J. R. Soc. Interface 9(71), 1318–1324 (2012).
Grunenfelder, L. K. et al. Biomineralization: Stress and damage mitigation from oriented nanostructures within the radular teeth of Cryptochiton stelleri. Adv. Funct. Mater. 24(39), 6093–6104 (2014).
Barber, A. H., Lu, D. & Pugno, N. M. Extreme strength observed in limpet teeth. J. R. Soc. Interface 12(105), 20141326 (2015).
Herrera, S. A., Grunenfelder, L., Escobar, E., Wang, Q., Salinas, C., Yaraghi, N., Geiger, J., Wuhrer, R., Zavattieri, P. & Kisailus, D. Stylus support structure and function of radular teeth. In Cryptochiton Stelleri, 20th International Conference on Composite Materials Copenhagen, 19–24th July, 2015.
Ukmar-Godec, T. et al. Materials nanoarchitecturing via cation-mediated protein assembly: Making limpet teeth without mineral. Adv. Mater. 29(27), 1701171 (2017).
Pohl, A. et al. Radular stylus of Cryptochiton stelleri: A multifunctional lightweight and flexible fiber-reinforced composite. J. Mech. Behav. Biomed. Mater. 111, 103991 (2020).
Stegbauer, L. et al. Persistent polyamorphism in the chiton tooth: From a new biomineral to inks for additive manufacturing. PNAS 118(23), e2020160118 (2021).
Weaver, J. C. et al. Analysis of an ultra hard magnetic biomineral in chiton radular teeth. Mater. Today 13(1–2), 42–52 (2010).
Wang, Q. et al. Phase transformations and structural developments in the radular teeth of Cryptochiton stelleri. Adv. Fun. Mater. 23, 2908–2917 (2013).
Ukmar-Godec, T. Mineralization of goethite in limpet radular teeth. In Iron Oxides: From Nature to Applications (eds Faivre, D. & Frankel, R. B.) 207–224 (Wiley-VCH, Weinheim, 2016).
Krings, W., Brütt, J.-O. & Gorb, S. N. Ontogeny of the elemental composition and the biomechanics of radular teeth in the chiton Lepidochitona cinerea. Under review at Frontiers in Zoology (2022).
Brooker, L. R. & Shaw, J. A. The chiton radula: A unique model for biomineralization studies. In Advanced Topics in Biomineralization (ed. Seto, J.) 65–84 (Intech Open, Rijeka, Croatia, 2012).
Joester, D. & Brooker, L. R. The chiton radula: A model system for versatile use of iron oxides. In Iron Oxides: From Nature to Applications (ed. Seto, J.) 177–205 (Wiley-VCH, Weinheim, 2016).
Kisailus, D. & Nemoto, M. Structural and proteomic analyses of iron oxide biomineralization in chiton teeth. In Biological Magnetic Materials and Applications (eds Matsunaga, T. et al.) 53–73 (Springer, Singapore, 2018).
Moura, H. M. & Unterlass, M. M. Biogenic metal oxides. Biomimetics 5(2), 29 (2020).
Krings, W., Kovalev, A., Glaubrecht, M. & Gorb, S. N. Differences in the Young modulus and hardness reflect different functions of teeth within the taenioglossan radula of gastropods. Zoology 137, 125713 (2019).
Krings, W., Neiber, M. T., Kovalev, A., Gorb, S. N. & Glaubrecht, M. Trophic specialisation reflected by radular tooth material properties in an ‘ancient’ Lake Tanganyikan gastropod species flock. BMC Ecol. Evol. 21, 35 (2021).
Krings, W., Marcé-Nogué, J. & Gorb, S. N. Finite element analysis relating shape, material properties, and dimensions of taenioglossan radular teeth with trophic specialisations in Paludomidae (Gastropoda). Sci. Rep. 11, 22775 (2021).
Gorb, S. N. & Krings, W. Mechanical property gradients of taenioglossan radular teeth are associated with specific function and ecological niche in Paludomidae (Gastropoda: Mollusca). Acta Biomater. 134, 513–530 (2021).
Troschel, F. H. Das Gebiss Der Schnecken Zur Begründung Einer Natürlichen Classification (Nicolaische Verlagsbuchhandlung, Berlin, Germany, 1863).
Sollas, I. B. The molluscan radula: Its chemical composition, and some points in its development. Q. J. Microsc. Sci. 51, 115–136 (1907).
Jones, E., McCance, R. & Shackleton, L. The role of iron and silica in the structure of the radular teeth of certain marine molluscs. J. Exp. Biol. 12(1), 59–64 (1935).
Tillier, S. & Cuif, J.-P. L’animal-conodonte est-il un Mollusque Aplacophore. C. R. Acad. Sci. Sér. 2 Méc. Phys. Chim. Sci. Univ. Sci. Terre 303(7), 627–632 (1986).
Cruz, R., Lins, U. & Farina, M. Minerals of the radular apparatus of Falcidens sp. (Caudofoveata) and the evolutionary implications for the phylum mollusca. Biol. Bull. 194(2), 224–230 (1998).
Smith, I. F. Lepidochitona cinerea, identification and biology, 2020. https://doi.org/10.13140/RG.2.2.28288.58889.
Smith, I. F. Acanthochitona fascicularis (Linnaeus, 1767), identification and biology, 2020. https://doi.org/10.13140/RG.2.2.10640.64005.
Quetglas, A., de Mesa, A., Ordines, F. & Grau, A. Life history of the deep-sea cephalopod family Histioteuthidae in the western Mediterranean. Deep Res. Part I Oceanogr. Res. Pap. 57, 999–1008 (2010).
Coelho, M., Domingues, P., Balguerias, E., Fernandez, M. & Andrade, J. P. A comparative study of the diet of Loligo vulgaris (Lamarck, 1799) (Mollusca: Cephalopoda) from the south coast of Portugal and the Saharan Bank (Central-East Atlantic). Fish. Res. 29(3), 245–255 (1997).
Notman, G. M., McGill, R. A., Hawkins, S. J. & Burrows, M. T. Macroalgae contribute to the diet of Patella vulgata from contrasting conditions of latitude and wave exposure in the UK. Mar. Ecol. Prog. Ser. 549, 113–123 (2016).
Marchais, V. et al. New tool to elucidate the diet of the ormer Haliotis tuberculata (L.): Digital shell color analysis. Mar. Biol. 164, 71 (2017).
Eichhorst, T. E. Neritidae of the World: Volume 1 and 2 (ConchBooks, 2016).
Bourguignat, M. J. R. Notice Prodromique sur les Mollusques Terrestres et Fluviatiles (Savy, Paris, 1885).
Bourguignat, M. J. R. Iconographie Malacologiques des Animaux Mollusques Fluviatiles du Lac Tanganika (Corbeil, Crété, 1888).
West, K., Michel, E., Todd, J., Brown, D. & Clabaugh, J. The gastropods of Lake Tanganyika: Diagnostic key, classification and notes on the fauna (Special publications: Societas Internationalis Limnologiae - Int. Assoc. of Theoretical and Applied Limnology, 2003)
Glaubrecht, M. Adaptive radiation of thalassoid gastropods in Lake Tanganyika, East Africa: Morphology and systematization of a paludomid species flock in an ancient lake. Zoosyst. Evol. 84, 71–122 (2008).
Moore, J. E. S. The Tanganyika Problem (Burst and Blackett, London, 1903).
Leloup, E. Exploration Hydrobiologique du Lac Tanganika (1946–1947) (Bruxelles, 1953).
Brown, D. Freshwater Snails of Africa and their Medical Importance (Taylor and Francis, London, 1994).
Germain, L. Mollusques du Lac Tanganyika et de ses environs. Extrait des resultats secientifiques des voyages en Afrique d’Edouard Foa. Bull. Mus. Natl. Hist. Nat. 14, 1–612 (1908).
Coulter, G. W. Lake Tanganyika and its Life (Oxford University Press, Oxford, 1991).
Bandel, K. Evolutionary history of East African fresh water gastropods interpreted from the fauna of Lake Tanganyika and Lake Malawi. Zent. Geol. Paläontol. Teil I, 233–292 (1997).
Pilsbry, H. A. & Bequaert, J. The aquatic mollusks of the Begian Congo. With a geographical and ecological account of Congo malacology. Bull. Am. Mus. Nat. Hist. 53, 69–602 (1927).
Lok, A. F. S. L., Ang, W. F., Ng, P. X., Ng, B. Y. Q. & Tan, S. K. Status and distribution of Faunus ater (Linnaeus, 1758) (Mollusca: Cerithioidea) in Singapore. NiS 4, 115–121 (2011).
Das, R. R. et al. Limited distribution of devil snail Faunus ater (Linnaeus, 1758) in tropical mangrove habitats of India. IJMS 47(10), 2002–2007 (2018).
Watson, D. C. & Norton, T. A. Dietary preferences of the common periwinkle, Littorina littorea (L.). J. Exp. Mar. Biol. Ecol. 88, 193–211 (1985).
Imrie, D. W., McCrohan, C. R. & Hawkins, S. J. Feeding behaviour in Littorina littorea: A study of the effects of ingestive conditioning and previous dietary history on food preference and rates of consumption. Hydrobiologia 193, 191–198 (1990).
Olsson, M., Svärdh, L. & Toth, G. B. Feeding behaviour in Littorina littorea: The red seaweed Osmundea ramosissima may not prevent trematode infection. Mar. Ecol. Prog. Ser. 348, 221–228 (2007).
Lauzon-Guay, J. S. & Scheibling, R. E. Food-dependent movement of periwinkles (Littorina littorea) associated with feeding fronts. J. Shellfish Res. 28, 581–587 (2009).
Bogan, A. E. & Hanneman, E. H. A carnivorous aquatic gastropod in the pet trade in North America: The next threat to freshwater gastropods?. Ellipsaria 15, 18–19 (2013).
Strong, E. E., Galindo, L. A. & Kantor, Y. I. Quid est Clea helena? Evidence for a previously unrecognized radiation of assassin snails (Gastropoda: Buccinoidea: Nassariidae). PeerJ 11(5), e3638 (2017).
Himmelman, J. H. & Hamel, J. R. Diet behaviour and reproduction of the whelk Buccinum undatum in the northern Gulf of St Lawrence, eastern Canada. Mar. Biol. 116, 423–430 (1993).
Barnes, H. & Powell, H. T. Onchidoris fusca (Müller); A predator of barnacles. J. Anim. Ecol. 23(2), 361–363 (1954).
Waters, V. L. Food-preference of the nudibranch Aeolidia papillosa, and the effect of the defenses of the prey on predation. Veliger 15(3), 174–192 (1973).
Edmunds, M., Potts, G., Swinfen, R. & Waters, V. The feeding preferences of Aeolidia papillosa (L.) (Mollusca, Nudibranchia). J. Mar. Biol. Assoc. U. K. 54(4), 939–947 (1974).
Edmunds, M. Advantages of food specificity in Aeolidia papillosa. J. Molluscan Stud. 49(1), 80–81 (1983).
Sørensen, C. G., Rauch, C., Pola, M. & Malaquias, M. A. E. Integrative taxonomy reveals a cryptic species of the nudibranch genus Polycera (Polyceridae) in European waters. J. Mar. Biol. Assoc. U. K. 100(5), 733–752 (2020).
Forrest, J. E. On the feeding habits and the morphology and mode of functioning of the alimentary canal in some littoral dorid nudibranchiate. Mollusca. Proc. Linn. Soc. Lond. 164(2), 225–235 (1953).
Rose, R. M. Functional morphology of the buccal mass of the nudibranch Archidoris pseudoargus. J. Zool. 165(3), 317–336 (1971).
Faivre, D. & Ukmar-Godec, T. From bacteria to mollusks: The principles underlying the biomineralization of iron oxide materials. Angew. Chem. Int. Ed. Engl. 54(16), 4728–4747 (2015).
Towe, K. M. & Lowenstam, H. A. Ultrastructure and development of iron mineralization in the radular teeth of Cryptochiton stelleri (Mollusca). J. Ultrastruct. Res. 17(1–2), 1–13 (1967).
Evans, L. A., Macey, D. J. & Webb, J. Distribution and composition of the matrix protein in the radula teeth of the chiton Acanthopleura hirtosa. Mar. Biol. 109, 281–286 (1991).
Macey, D. J. & Brooker, L. R. The junction zone: Initial site of mineralization in radula teeth of the chiton Cryptoplax striata (Mollusca: Polyplacophora). J. Morphol. 230, 33–42 (1996).
Lee, A. P. et al. In situ Raman spectroscopic studies of the teeth of the chiton Acanthopleura hirtosa. J. Biol. Inorg. Chem. 3, 614–619 (1998).
Brooker, L. R. & Macey, D. J. Biomineralization in chiton teeth and its usefulness as a taxonomic character in the genus Acanthopleura Guilding, 1829 (Mollusca: Polyplacophora). Am. Malacol. Bull. 16(1/2), 203–215 (2001).
Lee, A. P., Brooker, L. R., Macey, D. J., Webb, J. & van Bronswijk, W. A new biomineral identified in the cores of teeth from the chiton Plaxiphora albida. J. Biol. Inorg. Chem. 8(3), 256–262 (2003).
Shaw, J. A. et al. The chiton stylus canal: An element delivery pathway for tooth cusp biomineralization. J. Morphol. 270(5), 588–600 (2009).
Gordon, L. & Joester, D. Nanoscale chemical tomography of buried organic-inorganic interfaces in the chiton tooth. Nature 469, 194–198 (2011).
Emmanuel, S., Schuessler, J. A., Vinther, J., Matthews, A. & von Blanckenburg, F. A preliminary study of iron isotope fractionation in marine invertebrates (chitons, Mollusca) in near-shore environments. Biogeosciences 11(19), 5493–5502 (2014).
Shaw, J. A., Macey, D. J. & Brooker, L. R. Radula synthesis by three species of iron mineralizing molluscs: Production rate and elemental demand. J. Mar. Biol. Assoc. U. K. 88(3), 597–601 (2008).
Brooker, L. R., Lee, A. P., Macey, D. J., van Bronswijk, W. & Webb, J. Multiple-front iron-mineralisation in chiton teeth (Acanthopleura echinata: Mollusca: Polyplacophora). Mar. Biol. 142, 447–454 (2003).
Lee, A. P., Brooker, L. R., Macey, D. J., van Bronswijk, W. & Webb, J. Apatite mineralization in teeth of the chiton Acanthopleura echinata. Calcif. Tissue Int. 67, 408–415 (2000).
Brooker, L. R., Lee, A. P., Macey, D. J., Webb, J. & van Bronswijk, W. In situ studies of biomineral deposition in the radula teeth of chitons of the suborder Chitonina. Venus 65(1–2), 71–80 (2006).
van der Wal, P. Structure and formation of the magnetite-bearing cap of the polyplacophoran tricuspid radula teeth. In Iron Biominerals (eds Frankel, R. B. & Blakemore, R. P.) 221–229 (Plenum Press, New York, 1990).
Saunders, M., Kong, C., Shaw, J. A. & Clode, P. L. Matrix-mediated biomineralization in marine mollusks: A combined transmission electron microscopy and focused ion beam approach. Microsc. Microanal. 17, 220–225 (2011).
Lowenstam, H. A. Phosphatic hard tissues of marine invertebrates, their nature and mechanical function, and some fossil implications. Chem. Geol. 9, 153–166 (1972).
Macey, D. J., Webb, J. & Brooker, L. R. The structure and synthesis of biominerals in chiton teeth. Bull. Inst. Océanogr. (Monaco) 4(1), 191–197 (1994).
Lowenstam, H. A. & Weiner, S. Transformation of amorphous calcium phosphate to crystalline dahllite in the radula teeth of chitons. Science 227, 51–52 (1985).
Lowenstam, H. A. & Weiner, S. Mollusca. In On biomineralization (eds Lowenstam, H. A. & Weiner, S.) 88–305 (Oxford University Press, Oxford, 1989).
Evans, L. A. & Alvarez, R. Characterization of the calcium biomineral in the radular teeth of Chiton pelliserpentis. J. Biol. Inorg. Chem. 4(2), 166–170 (1999).
Evans, L. A., Macey, D. J. & Webb, J. Calcium biomineralization in the radula teeth of the chiton, Acanthopleura hirtosa. Calcif. Tissue Int. 51, 78–82 (1992).
Kim, K. S., Webb, J., Macey, D. J. & Cohen, D. D. Compositional changes during biomineralization of the radula of the chiton Clavarizona hirtosa. J. Inorg. Biochem. 28(2–3), 337–345 (1986).
Runham, N. W. The histochemistry of the radula of Patella vulgata. Q. J. Microsc. Sci. 102(3), 371–380 (1961).
Runham, N. W., Thronton, P. R., Shaw, D. A. & Wayte, R. C. The mineralization and hardness of the radular teeth of the limpet Patella vulgate L. Z. Zellforsch. 99, 608–626 (1969).
Grime, G. et al. Biological applications of the Oxford scanning proton microprobe. Trends Biochem. Sci. 10(1), 6–10 (1985).
St Pierre, T. G. et al. Iron oxide biomineralization in the radula teeth of the limpet Patella vulgata; Mössbauer spectroscopy and high resolution transmission electron microscopy studies. Proc. R. Soc. B 228, 31–42 (1986).
Mann, S., Perry, C. C., Webb, J., Luke, B. & Williams, R. J. P. Structure, morphology, composition and organization of biogenic minerals in limpet teeth. Proc. R. Soc. B 227(1247), 179–190 (1986).
van der Wal, P. Structural and material design of mature mineralized radula teeth of Patella vulgata (Gastropoda). J. Ultrastruct. Mol. Struct. Res. 102(2), 147–161 (1989).
Huang, C., Li, C.-W., Deng, M. & Chin, T. Magnetic properties of goethite in radulae of limpets. IEEE Trans. Magn. 28(5), 2409–2411 (1992).
Rinkevich, B. Major primary stages of biomineralization in radular teeth of the limpet Lottia gigantea. Mar. Biol. 117, 269–277 (1993).
Liddiard, K. J., Hockridge, J. G., Macey, D. J., Webb, J. & van Bronswijk, W. Mineralisation in the teeth of the limpets Patelloida alticostata and Scutellastra laticostata (Mollusca: Patellogastropoda). Molluscan Res. 24, 21–31 (2004).
Cruz, R. & Farina, M. Mineralization of major lateral teeth in the radula of a deep-sea hydrothermal vent limpet (Gastropoda: Neolepetopsidae). Mar. Biol. 147, 163–168 (2005).
Davies, M. S., Proudlock, D. J. & Mistry, A. Metal concentrations in the radula of the common limpet, Patella vulgata L., from 10 sites in the UK. Ecotoxicology 14(4), 465–475 (2005).
Sone, E. D., Weiner, S. & Addadi, L. Biomineralization of limpet teeth: A cryo-TEM study of the organic matrix and the onset of mineral deposition. J. Struct. Biol. 158, 428–444 (2007).
Hua, T.-E. & Li, C.-W. Silica biomineralization in the radula of a limpet Notoacmea schrenckii (Gastropoda: Acmaeidae). Zool. Stud. 46(4), 379–388 (2007).
Krings, W. et al. In slow motion: Radula motion pattern and forces exerted to the substrate in the land snail Cornu aspersum (Mollusca, Gastropoda) during feeding. R. Soc. Open Sci. 6(7), 2054–5703 (2019).
Mikovari, A. et al. Radula development in the giant key-hole limpet Megathura crenulate. J. Shellfish Res. 34(3), 893–902 (2015).
Ukmar-Godec, T., Kapun, G., Zaslansky, P. & Faivre, D. The giant keyhole limpet radular teeth: A naturally-grown harvest machine. J. Struct. Biol. 192, 392–402 (2015).
Macey, D. J., Brooker, L. R. & Cameron, V. Mineralisation in the teeth of the gastropod mollusc Nerita atramentosa. Molluscan Res. 18(1), 33–41 (1997).
Barkalova, V. O., Fedosov, A. E. & Kantor, Y. I. Morphology of the anterior digestive system of tonnoideans (Gastropoda: Caenogastropoda) with an emphasis on the foregut glands. Molluscan Res. 36, 54–73 (2016).
Ponte, G. & Modica, M. V. Salivary glands in predatory mollusks: Evolutionary considerations. Front. Physiol. 8, 580 (2017).
Haszprunar, G. On the origin and evolution of major gastropod groups, with special reference to the Streptoneura. J. Molluscan Stud. 54, 367–441 (1988).
Sasaki, T. Comparative anatomy and phylogeny of the recent Archaeogastropoda (Mollusca: Gastropoda). Univ. Tokyo Bull. 38, 1–224 (1998).
Simone, L. R. L. Phylogeny of the Caenogastropoda (Mollusca), based on comparative morphology. Arq. Zool. 42(4), 161–323 (2011).
Meirelles, C. A. & Matthews-Cascon, H. Relations between shell size and radula size in marine prosobranchs (Mollusca: Gastropoda). Thalassas 19(2), 45–53 (2003).
Peile, A. J. Some radula problems. J. Conchol. 20, 292–304 (1937).
Marcus, E. & Marcus, E. Mesogastropoden von der Küste São Paulos. Abh Math Naturwissenschaftlichen Kl Akad Wiss Lit Mainz 1963(1), 1–105 (1963).
Reid, D. G. The Littorinid Molluscs of Mangrove Forests in the Indo-Pacific Region: The Genus LITTORARIA (British Museum Natural History, London, 1986).
Reid, D. G. The comparative morphology, phylogeny and evolution of the gastropod family Littorinidae. Philos. Trans. R. Soc. Lond. B 324, 1–110 (1989).
Reid, D. G. & Mak, Y.-M. Indirect evidence for ecophenotypic plasticity in radular dentition of Littoraria species (Gastropoda: Littorinidae). J. Molluscan Stud. 65(3), 355–370 (1999).
Fretter, V. & Graham, A. British Prosobranch Molluscs (The Ray Society, London, 1994).
Cabral, J. P. Shape and growth in European Atlantic Patella limpets (Gastropoda, Mollusca). Ecological implications for survival. Web Ecol. 7, 11–21 (2007).
Nesson, M. H. Studies on radula tooth mineralization in the Polyplacophora, thesis, California Institute of Technology, Pasadena, USA (1969).
Shaw, J. A., Brooker, L. R. & Macey, D. J. Radular tooth turnover in the chiton Acanthopleura hirtosa (Blainville, 1825) (Mollusca: Polyplacophora). Molluscan Res. 22, 93–99 (2002).
Isarankura, K. & Runham, N. Studies on the replacement of the gastropod radula. Malacologia 7(1), 71–91 (1968).
Padilla, D. K., Dittman, D. E., Franz, J. & Sladek, R. Radular production rates in two species of Lacuna Turton (Gastropoda: Littorinidae). J. Molluscan Stud. 62(3), 275–280 (1996).
Runham, N. W. Rate of replacement of the molluscan radula. Nature 194, 992–993 (1962).
Mackenstedt, U. & Märkel, K. Experimental and comparative morphology of radula renewal in pulmonates (Mollusca, Gastropoda). Zoomorphology 107(4), 209–239 (1987).
Mischor, B. & Märkel, K. Histology and regeneration of the radula of Pomacea bridgesi (Gastropoda, Prosobranchia). Zoomorphology 104, 42–66 (1984).
Fujioka, Y. Seasonal aberrant radular formation in Thais bronni (Dunker) and T. clavigera (Küster) (Gastropoda: Muricidae). J. Exp. Mar. Biol. Ecol. 90(1), 43–54 (1985).
Liu, Z., Meyers, M. A., Zhang, Z. & Ritchie, R. O. Functional gradients and heterogeneities in biological materials: Design principles, functions, and bioinspired applications. Progr. Mater. Sci. 88, 467–498 (2017).
Vincent, J. F. V. The hardness of the tooth of Patella vulgata L. Radula: A Reappraisal. J. Molluscan Stud. 46, 129–133 (1980).
Evans, L. A., Macey, D. J. & Webb, J. Characterization and structural organization of the organic matrix of radula teeth of the chiton Acanthopleura hirtosa. Philos. Trans. R. Soc. Lond. B 329, 87–96 (1990).
Evans, L. A., Macey, D. J. & Webb, J. Matrix heterogeneity in the radular teeth of the chiton Acanthopleura hirtosa. Acta Zool. 75(1), 75–79 (1994).
Wealthall, R. J., Brooker, L. R., Macey, D. J. & Griffin, B. J. Fine structure of the mineralized teeth of the chiton Acanthopleura echinata (Mollusca: Polyplacophora). J. Morphol. 265, 165–175 (2005).
Krings, W., Kovalev, A. & Gorb, S. N. Influence of water content on mechanical behaviour of gastropod taenioglossan radulae. Proc. R. Soc. B 288, 20203173 (2021).
Krings, W., Kovalev, A. & Gorb, S. N. Collective effect of damage prevention in taenioglossan radular teeth is related to the ecological niche in Paludomidae (Gastropoda: Cerithioidea). Acta Biomater. 135, 458–472 (2021).
Radwin, G. E. & Wells, H. W. Comparative radular morphology and feeding habits of muricid gastropods from the Gulf of Mexico. Bull. Mar. Sci. 18(1), 72–85 (1968).
Grünbaum, D. & Padilla, D. K. An integrated modeling approach to assessing linkages between environment, organism, and phenotypic plasticity. Integr. Comp. Biol. 54(2), 323–335 (2014).
Scheel, C., Gorb, S. N., Glaubrecht, M. & Krings, W. Not just scratching the surface: Distinct radular motion patterns in Mollusca. Biol. Open 9, bio055699 (2020).
Gray, J. On the division of ctenobranchous gasteropodous Mollusca into larger groups and families. Ann. Mag. Nat. Hist. 11(2), 124–133 (1853).
Hyman, L. H. Mollusca I. Aplacophora polyplacophora monoplacophora. Gastropoda, the coelomate bilateria. The invertebrates 6 (McGraw-Hill Book Company, New York, 1967).
Nixon, M. A nomenclature for the radula of the Cephalopoda (Mollusca) – living and fossil. J. Zool. 236, 73–81 (1995).
Haszprunar, G. & Götting, K. J. Mollusca, Weichtiere. In Spezielle Zoologie Teil Einzeller und wirbellose Tiere (eds Westheide, W. & Rieger, R.) 305–362 (Springer, Berlin, Germany, 2007).
Lowenstam, H. A. Magnetite in denticle capping in recent chitons (Polyplacophora). Geol. Soc. Am. Bull. 73, 435–438 (1962).
Kirschvink, J. L. & Lowenstam, H. A. Mineralization and magnetization of chiton teeth: Paleomagnetic, sedimentalogic and biologic implications of organic magnetite. EPSL 44, 193–204 (1979).
Han, Y. et al. Magnetic and structural properties of magnetite in radular teeth of chiton Acanthochiton rubrolinestus. Bioelectromagnetics 32, 226–233 (2011).
Nemoto, M. et al. Integrated transcriptomic and proteomic analyses of a molecular mechanism of radular teeth biomineralization in Cryptochiton stelleri. Sci. Rep. 9, 856 (2019).
McCoey, J. M. et al. Quantum magnetic imaging of iron biomineralization in teeth of the chiton Acanthopleura hirtosa. Small Methods 4, 1900754 (2020).
Lowenstam, H. A. Lepidocrocite, an apatite mineral, and magnetite in teeth of chitons (Polyplacophora). Science 56, 1373–1375 (1967).
Brooker, L. R., Lee, A. P., Macey, D. J. & Webb, J. Molluscan and other marine teeth. In Encyclopedia of Materials: Science and Technology (eds Buschow, K. H. J. et al.) 5186–5189 (Elsevier Science Ltd., Oxford, 2001).
Shaw, J. A. et al. Ultrastructure of the epithelial cells associated with tooth biomineralization in the chiton Acanthopleura hirtosa. Microsc. Microanal. 15(2), 154–165 (2009).
Creighton, T. E. Protein folding coupled to disulphide bond formation. Biol. Chem. 378(8), 731–744 (1997).
Harding, M. M. Metal-ligand geometry relevant to proteins and in proteins: Sodium and potassium. Acta Cryst. D 58, 872–874 (2002).
Hayes, T. The influence of diet on local distributions of Cypraea. Pac. Sci. 37(1), 27–36 (1983).
Padilla, D. K. The importance of form: Differences in competitive ability, resistance to consumers and environmental stress in an assemblage of coralline algae. J. Exp. Mar. Biol. Ecol. 79(2), 105–127 (1984).
Kesler, D. H., Jokinen, E. H. & Munns, W. R. Jr. Trophic preferences and feeding morphology of two pulmonate snail species from a small New England pond, USA. Can. J. Zool. 64, 2570–2575 (1986).
Blinn, W., Truitt, R. E. & Pickart, A. Feeding ecology and radular morphology of the freshwater limpet Ferrissia fragilis. J. N. Am. Benthol. Soc. 8(3), 237–242 (1989).
Hawkins, S. J. et al. A comparison of feeding mechanisms in microphagous, herbivorous, intertidal, prosobranchs in relation to resource partitioning. J. Molluscan Stud. 55(2), 151–165 (1989).
Franz, C. J. Feeding patterns of Fissurella species on Isla de Margarita, Venezuela: Use of radulae and food passage rates. J. Molluscan Stud. 56(1), 25–35 (1990).
Thompson, R. C., Johnson, L. E. & Hawkins, S. J. A method for spatial and temporal assessment of gastropod grazing intensity in the field: The use of radula scrapes on wax surfaces. J. Exp. Mar. Biol. Ecol. 218(1), 63–76 (1997).
Iken, K. Feeding ecology of the Antarctic herbivorous gastropod Laevilacunaria antarctica Martens. J. Exp. Mar. Biol. Ecol. 236(1), 133–148 (1999).
Forrest, R. E., Chapman, M. G. & Underwood, A. J. Quantification of radular marks as a method for estimating grazing of intertidal gastropods on rocky shores. J. Exp. Mar. Biol. Ecol. 258(2), 155–171 (2001).
Dimitriadis, V. K. Structure and function of the digestive system in Stylommatophora. In The Biology of Terrestrial Molluscs (ed. Barker, G. M.) 237–258 (CABI Publishing, Wallingford, UK, 2001).
Speiser, B. Food and feeding behaviour. In The Biology of Terrestrial Molluscs (ed. Barker, G. M.) 259–288 (CABI Publishing, Wallingford, UK, 2001).
Sitnikova, T. et al. Resource partitioning in endemic species of Baikal gastropods indicated by gut contents, stable isotopes and radular morphology. Hydrobiologia 682, 75–90 (2012).
Bergmeier, F. S., Ostermair, L. & Jörger, K. M. Specialized predation by deep-sea Solenogastres revealed by sequencing of gut contents. Curr. Biol. 31(13), R836–R837 (2021).
Goodheart, J. A., Bazinet, A. L., Valdés, Á., Collins, A. G. & Cummings, M. P. Prey preference follows phylogeny: Evolutionary dietary patterns within the marine gastropod group Cladobranchia (Gastropoda: Heterobranchia: Nudibranchia). BMC Evol. Biol. 17, 221 (2017).
Padilla, D. K. Structural resistance of algae to herbivores. A biomechanical approach. Mar. Biol. 90, 103–109 (1985).
Padilla, D. K. Algal structural defenses: Form and calcification in resistance to tropical limpets. Ecology 70(4), 835–842 (1989).
Wilson, A. B., Glaubrecht, M. & Meyer, A. Ancient lakes as evolutionary reservoirs: Evidence from the thalassoid gastropods of Lake Tanganyika. Proc. R. Soc. B 271(1538), 529–536 (2004).
Ponder, W. & Lindberg, D. R. Phylogeny and Evolution of the Mollusca (University of California Press, Berkeley, California, 2008).
Jörger, K. M. et al. On the origin of Acochlidia and other enigmatic euthyneuran gastropods, with implications for the systematics of Heterobranchia. BMC Evol. Biol. 10, 323 (2010).
Kocot, K. et al. Phylogenomics reveals deep molluscan relationships. Nature 477, 452–456 (2011).
Kocot, K. M., Poustka, A. J., Stöger, I., Halanych, K. M. & Schrödl, M. New data from Monoplacophora and a carefully-curated dataset resolve molluscan relationships. Sci. Rep. 10, 101 (2020).
Smith, S. et al. Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature 480, 364–367 (2011).
Haszprunar, G. & Wanninger, A. Molluscs. Curr Biol. 22(13), R510-514 (2012).
Wanninger, A. & Wollesen, T. The evolution of molluscs. Biol. Rev. 94, 102–115 (2019).
Irisarri, I., Uribe, J. E., Eernisse, D. J. & Zardoya, R. A mitogenomic phylogeny of chitons (Mollusca: Polyplacophora). BMC Evol. Biol. 20, 22 (2020).
Acknowledgements
We would like to thank Renate Walter and Elke Woelken from the Zoological Institute, Universität Hamburg (UHH), Germany, for their support with the SEM and Thomas Schwaha from the Universität Wien, Austria, for identifying specimens of Acanthochitona fascicularis and providing their habitus images. We are grateful for the help of the members from the department “Functional Morphology and Biomechanics”, Christian-Albrechts-Universität zu Kiel (CAU), Germany, for collecting individuals of Lepidochitona cinerea and Littorina littorea at Husum during the annual work outing, Marco T. Neiber from the UHH for his support on species identification, and Thomas M. Kaiser from the Leibniz Institute for the Analysis of Biodiversity Change (LIB), Hamburg, Germany, for some discussion on the results. We thank Frank Friedrich from the Zoological Institute (UHH) for supervising the master thesis of OB, serving as basis for this study, and his advices on EDX data interpretation, Alexandra Kerbl from the University of Exeter, UK, and Lenke Tödter from the LIB for collecting individuals of Patella vulgata, Ute Kieb from the Alfred-Wegener-Institute from Helgoland for providing specimens of Buccinum and individuals of Aeolidia, Onchidoris, Polycera, and Doris. We are grateful for the access to Ilka Sötje’s highly equipped lab at the UHH, which enabled us to keep and feed the nudibranch molluscs, and her expertise on the preservation of marine invertebrates, the lecturers of the students’ dissection course from the UHH and LIB for reserving Loligo’s radulae and beaks, Heinz Büscher from Basel, Switzerland, for collecting specimens of the Paludomidae at Lake Tanganyika, and Bernhard Hausdorf from the LIB for providing access to Histioteuthis specimens. We are grateful for the technical support of Esther Appel and Alexander Kovalev from the Department “Functional Morphology and Biomechanics” (CAU) at the beginning of the study, and the helpful comments of the two anonymous reviewers.
Funding
Open Access funding enabled and organized by Projekt DEAL. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
W.K. and S.G. together initiated the project, designed the study, and discussed the data. In the course of his master thesis, J.O.B. performed EDX analyses of six species and collated the literature. WK together with J.O.B. wrote the first draft of the manuscript. W.K. executed the remaining EDX anal yses and created the figures. All authors contributed to and approved the final version of the manuscript for publication.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Krings, W., Brütt, JO. & Gorb, S.N. Elemental analyses reveal distinct mineralization patterns in radular teeth of various molluscan taxa. Sci Rep 12, 7499 (2022). https://doi.org/10.1038/s41598-022-11026-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41598-022-11026-w
This article is cited by
Mandible mechanical properties and composition of the larval Glossosoma boltoni (Trichoptera, Insecta)
Scientific Reports (2024)
Mechanical properties, degree of sclerotisation and elemental composition of the gastric mill in the red swamp crayfish Procambarus clarkii (Decapoda, Crustacea)
Scientific Reports (2022)
The ontogeny of elements: distinct ontogenetic patterns in the radular tooth mineralization of gastropods
The Science of Nature (2022)
Material gradients in gastropod radulae and their biomechanical significance: a combined approach on the paludomid Lavigeria grandis
The Science of Nature (2022)
Comments
By submitting a comment you agree to abide by our Terms and Community Guidelines. If you find something abusive or that does not comply with our terms or guidelines please flag it as inappropriate.
