
Abstract
Biological control and taxonomy are continuously developing fields with remarkable impacts on society. At least 80 years of literature have documented this relationship, which remains essentially the same in its mutualistic nature, as well as in its major challenges. From the perspective of Brazilian taxonomists, we discuss the impacts of important scientific and social developments that directly affect research in these areas, posing new challenges for this lasting relationship. The increasing restrictions and concerns regarding the international transit of organisms require improvements in research related to risk assessment for exotic biological control agents and also stimulate prospecting within the native biota. In our view, this is a positive situation that can foster a closer relationship between taxonomists and applied entomologists, as well as local surveys and taxonomic studies that are necessary before new programs and agents can be implemented. We discuss the essential role of molecular biology in this context, as an iconic example of the synergy between applied sciences and natural history. As our society comes to need safer and more sustainable solutions for food security and the biodiversity crisis, scientific progress will build upon this integration, where biological control and taxonomy play an essential role.
Similar content being viewed by others


Perspectives and overview
Since the early days of biological control as a scientific field, its close relationship with taxonomy has remained essentially unchanged. One field has fostered the development of the other, leading to advances in the recognition of species and knowledge of their biology, behavior, and evolutionary and ecological relationships. The study of natural enemies of pests, such as parasitoids, has benefited from this integration, even though much work remains to be done. Recent examples show that even in critical situations, whether from an economic or conservation perspective, highly important groups for biological control programs still lack adequate taxonomic support. Thus, the knowledge, investments, and actions taken may not only be completely lost but also generate negative results (Bortolus 2008). The same problems and possible solutions raised by authors in the past remain relevant today, as unexplored problems and possibilities.
The science of taxonomy has biological diversity as its object of study, describing and classifying organisms using the comparison of characters between distinct evolutionary entities, or taxa (i.e., base units of taxonomy including species and higher-level groups such as genus and family) as a method (Amorim 1997). Taxonomic work extends beyond species description and identification in that it often includes collections and surveys, maintenance of biological collections and museums, composition of catalogs and databases, and phylogenetic studies, which can provide important information about evolutionary aspects such as geographic distribution and host relationships. It is a hypothesis-driven science, in which taxa are proposed and tested in a complex routine. Therefore, a new species description involves the generation of a new hypothesis represented by a name, which in turn is governed by nomenclature rules (Thiele and Yeates 2002; Lipscomb et al. 2003; Wheeler 2004; Thomson et al. 2018). For the purposes of the present discussion, we consider three levels of taxonomic studies: alpha (species level, their discovery and description), beta (supraspecific level, phylogenetic relationships and classification), and gamma (infraspecific level and speciation processes) (Mayr 1958). We use the terms “taxonomy” and “systematics” interchangeably, as we see no advantage in divorcing these terms (Wheeler 2007) or a logical difference between them (see Hennig 1999).
Biological control, in turn, is a branch of applied ecology, which derives most of its theoretical basis from population ecology and demography in a consumer-resource dynamic (Murdoch et al. 2003). The results are usually integrated into environmental management practices, aiming to reduce undesirable populations which may be agricultural or forest pests, urban pests, or invasive species in natural areas. Over the years, several books and articles have provided more general explanations about the applicable terminology, such as DeBach and Rosen (1991), Van Driesche and Bellows (1996), Eilenberg et al. (2001), and Parra et al. (2002; 2021). Biological control programs may have positive impacts on food security and the quality of agricultural products or the recovery of natural areas, but may also lead to population reductions and even extinction of native species. These potential outcomes place great responsibility on researchers and practitioners, which increases the need for taxonomic and ecological studies of the species involved (Van Driesche et al. 2016).
The different methods of biological control (see Eilenberg et al. 2001, for a discussion of terminology applied to biological control strategies) vary in their potential effects on the environment and the economics of the productive sector. For example, classical biological control aims at establishing populations of a biological control agent and its subsequent dispersal in the new environment. This approach can produce irreversible results, through both target control and non-target effects (Howarth 1991). It is not directed at mass production and marketing of the agent, but benefits producers in a broad and general way (e.g., Neuenschwander 2001) and can help to mitigate the environmental impacts of invasive species (Van Driesche et al. 2016). Applied biological control, including inundationFootnote 1 and inoculation, depends on mass rearing and release of agents, and therefore on methods to produce and apply the product (i.e., the agent); a review of the state of this area was published by van Lenteren (2012). In most cases, the establishment of the agent after release is not a target attribute of this approach, and its impact, positive or negative, tends to be transient (Boivin et al. 2006). Finally, conservation biological control depends exclusively on native fauna and tends to generate additional benefits, with an increase in local biodiversity (Barbosa 1998). This method is also referred to as “ecological engineering” (Gurr et al. 2004) and is based on habitat manipulation, aiming to provide resources for natural enemies to improve natural control in agroecosystems (Altieri and Letourneau 1982).
In all these cases, knowledge and correct determination of the species involved is a fundamental step. In addition, because there can be significant biological differences between populations of a species, biological control often requires gamma-level taxonomic studies, that is, what has been referred to as biosystematics (Schlinger and Doutt 1964; Gordh 1977). Many advances in this area were also responses to the demands of biological control programs, including proposals in integrative taxonomy (for example, Dayrat 2005; Schlick-Steiner et al. 2010; Beltrà et al. 2015; Gokhman 2018). Taxonomically important research studies involving species concepts, cryptic species, and hybridization (for example, Goldson et al. 2003; Hopper et al. 2006; Bickford et al. 2007; Yara et al. 2010; Derocles et al. 2016) have been stimulated thanks to their relevance to biological control.
From Clausen (1942) to Zucchi (2021), a vast literature extending over at least eight decades has discussed the intricacies and the intimate mutualistic relationship between taxonomic and biological control research (Clausen 1942; Sabrosky 1955; DeBach 1960; Schlinger and Doutt 1964; Compere 1969; Rosen and DeBach 1973; Delucchi et al. 1976; Gordh 1977; Knutson 1981; Rosen 1986; Moraes 1987; Danks 1988; Hanson 1993; Heraty 1998, 2004, 2009; Schauff and LaSalle 1998; Gordh and Beardsley 1999; Huber et al. 2002, 2021; Zucchi 2002, 2021; González-Hernández and López-Arrollo 2007; Bin et al. 2012; Andersen and Wagner 2016). Despite the exhaustive discussion of this topic, which undeniably denotes its importance, the desirable cooperation between the fields of taxonomy and biological control still occurs only in particular cases. To some degree, the rhetorical question raised by Sabrosky (1955) regarding taxonomic knowledge of an agent still seems to echo in practice: “What difference does it make, if control is achieved?” This is a valid question, since the entities in question, whether insect pests or biological control agents, have characteristics that allow them to be distinguished as important pests or efficient agents, regardless of the names assigned to them. In the taxonomist’s search for natural groups or for a more appropriate philosophical concept of species, the name could not be a more artificial aspect. Could this realization cause a rupture between the two fields of knowledge? For most authors who have discussed the topic, scientific communication justifies the importance of taxonomy. However, the distinction between real entities and abstract concepts of species and populations must be explicit, allowing unambiguous understanding by the researchers involved. To this end, some situations may require an infraspecific nomenclatural system (Kim and McPheron 1993).
Even in the best possible scenario, an identification may still be flawed and the name assigned to a particular entity may not be the most accurate, or the species concept not appropriate, or even the samples used may originate from mixed populations that differ in their biology. History shows that all these problems, among many others, have already occurred, which highlights the need for careful, in-depth studies of both taxonomic and ecological aspects, as flawed identifications can lead to large economic losses, lack of confidence in biological control, and environmental impacts (e.g., Louda et al. 2003; Bin et al. 2012). Perhaps the most important point is understanding that taxonomic issues will occasionally occur, as taxonomic concepts are neither universal nor stable (Minelli 2020), and that some actions, such as introduction of agents into new areas, are irreversible. For both problems, there are best practices that must be followed by both taxonomists and applied entomologists, and can increase efficiency and safety and, consequently, confidence in biological control as a management practice, through closer interaction between these two fields of research.
With the recent developments in these fields, increased specialization seems to be the trend. In the past, scientists such as David Rosen and Paul DeBach were able to conduct research in both taxonomy and biological control. Today, they would probably encounter more difficulty in conducting studies in both areas. How this change may affect this collaborative relationship is not yet clear. This opinion paper aims to explore aspects of the relationship between insect taxonomy, especially parasitoids, and biological control, that affect the development of these areas and their mutualistic relationship.
Taxonomy in the context of biological control
Three fundamental aspects of biological control increase the importance of taxonomy. First, control agents are species (or a lineage within a species), and therefore need to be studied taxonomically: species must be discovered/delimited, described, and named within a classification based on their phylogenetic (= evolutionary) relationships, and this process must ensure accurate identification of the species in the future. Second, it is not uncommon for biological control agents to be more specific than chemical agents, making correct identification of the target even more crucial to success. A third, emerging factor is the association between the pest and the natural enemy. Identification and observations of these associations are not infrequently subject to errors that cause numerous problems in biological control programs (Danks 1988; Bin et al. 2012).
Taxonomy of insects, especially natural enemies of pests, has certain aspects that should be considered for a better understanding of the relationship to biological control: (i) there is an immense diversity of insects (mostly unknown) and species delimitation is often, if not always, a complex task; (ii) biological control requires a fine delimitation of species, and sometimes infraspecific refinement of strains or populations within a species, which has led taxonomists to push the boundaries of alpha and gamma levels; (iii) taxonomists are aware of the need for identification tools for use by applied entomologists, and the development of such tools may increase identification accuracy and expand the use of biological control agents, even in the most complex groups; (iv) the amount of information on species of economic importance is, as expected, much larger than on other species with no direct/obvious importance to humans, and so are the cumulative errors in identification, especially in host-parasitoid associations; (v) molecular markers are rapidly increasing in importance in taxonomy, and research should also focus on curating public databases; however, in many cases, published information is impossible to confirm due to a lack of voucher specimens (Packer et al. 2018).
The diversity of insects is the highest among living beings, although about 80% of species remain to be described (Stork 2018). Hymenopteran parasitoids, which are arguably the most important group of macroorganisms in biological control, are an excellent example of the scale of this knowledge gap. According to Forbes et al. (2018), the number of unknown species of parasitoid wasps must be around twice that of the order Coleoptera, currently the group with the largest number of described species. These numbers are based on estimates, such as that of Hawkins and Lawton (1987), that a typical phytophagous species is attacked by 5–10 species of parasitoids. Besides the evident lack of taxonomic studies, this immense diversity may be partly masked by the high frequency of cryptic species and minute size of these organisms (Gokhman 2018), exacerbating the complexity of species delimitation and identification. In addition, many environments have been scarcely studied, which makes it difficult to estimate the true diversity of this group of wasps (Grissell 1999; Sharkey and Fernández 2006; Forbes et al. 2018; Stork 2018).
Despite this deficit in knowledge, economically important groups are relatively well studied, such as the chalcidoids (Hymenoptera) Trichogramma Westwood (e.g., Pinto 1999; Borghuis et al. 2004; Zucchi et al. 2010; Parra and Coelho 2019; Querino and Zucchi 2019), Encarsia Foerster (e.g., Heraty and Polaszek 2000; Manzari et al. 2002; Gebiola et al. 2016a, 2016b), and Gonatocerus Nees (de León et al. 2004, 2006; Vickerman et al. 2004; Triapitsyn 2006; Triapitsyn et al. 2010; Huber 2015). These taxonomic studies have clarified the identity of the species, often based exclusively on morphological characters, regardless of the minute body size of these wasps. Likewise, geographic distributions are better known for species of economic importance. For Braconidae (Hymenoptera), for instance, the two species with the most records in Brazil (Yu et al. 2016), Doryctobracon areolatus (Szépligeti) and Opius bellus Gahan, are both important in the control of several tephritid fruit flies (Marinho et al. 2018).
Well-studied groups tend to have relatively more synonyms, as taxonomists may have different concepts or may fail to effectively communicate these concepts (Gaston and Mound 1993). A negative effect of focusing on species of economic importance is reflected in the large number of synonyms. For example, the species of aphids with the most synonyms listed in the catalog by Remaudière and Remaudière (1997) are widespread polyphagous agricultural pests, including Aphis gossypii Glover, Aphis fabae Scopoli, Aulacorthum solani (Kaltenbach), Brachycaudus helichrysi (Kaltenbach), and Myzus persicae (Sulzer), with more than 30 and up to 47 junior synonyms (Miller and Foottit 2009). Besides polyphagy, morphological variations on different hosts, and difficulties in communication among researchers, including access to type material (Hunter 1901; Ilharco and van Harten 1987), economic interest is largely responsible for these high numbers. These long lists of synonyms may obscure species identification, due to the concatenation of all associated ecological records and other aspects, or hide relevant intraspecific variation that can only be revealed by an integrative approach, as done for B. helichrysi for example (Piffaretti et al. 2012).
The same is true for parasitoids (see the example of the Peristenus pallipes complex in the section “Recent examples”); and for associations, the issue reaches another level of complexity. Noyes (1994) reviewed the reliability of host-parasitoid association data and demonstrated that most records, including many for well-known species such as Cotesia glomerata (L.) (Hymenoptera, Braconidae) and its hosts, or Plutella xylostella (L.) (Lepidoptera, Plutellidae) and its natural enemies, were in fact incorrect. For a parasitoid widely used in biological control such as Lysiphlebus testaceipes (Cresson) (Hymenoptera, Braconidae), with many known synonyms (21) and dozens of recorded hosts, including A. gossypii and A. fabae (Yu et al. 2016), it is highly probable that a significant portion of reported associations are observation errors (Bin et al. 2012). One of the most important issues in biological control that arises from the existence of polyphagy as well as large numbers of cryptic species and synonyms was examined by Derocles et al. (2016) in the article “Are generalist Aphidiinae (Hym. Braconidae) mostly cryptic species complexes?” The frequency of such cases in economically important species makes catalogs even more important, especially catalogs for natural enemies that include their host associations, such as those for Ichneumonoidea (Yu et al. 2016) and Chalcidoidea (Noyes 2019).
One final issue involving taxonomy and economically important species (i.e., pests and their natural enemies) is their higher likelihood of crossing natural barriers and colonize areas outside their original distribution aided by human action. This poses a challenge for taxonomists, especially when the species in question belongs to a group (e.g., a genus) with a cosmopolitan distribution and an inadequately documented introduction history. Species might be described for the first time from individuals that formed populations outside their native range, as in the case of the Chinese mealybug Ceroplastes sinensis Del Guercio (Hemiptera, Coccidae), an important citrus pest with worldwide distribution, eventually discovered to be native to Argentina (Qin et al. 1994). Similarly, the important orchid pest Contarinia maculipennis Felt (Diptera, Cecidomyiidae) was first described in Hawai’i but later found to be native to Southeast Asia (Gagné 1995; Uechi et al. 2003). For cosmopolitan species such as Pachycrepoideus vindemmiae (Rondani) (Hymenoptera, Pteromalidae), the center of origin and how they spread across the planet remain unknown (Shimbori et al. 2020). Obviously, such situations may hinder any search for biological control agents within the native range of the target pest. Furthermore, the recent profound changes in international transit of species through human actions, including the intentional introduction of biological control agents, require even greater collaboration among taxonomists, applied entomologists, and regulators, as discussed below.
Changes in biological control, the role of regulations, and impacts on taxonomy
The advent of stronger restrictions on the introduction of exotic biological control agents around the world has changed the biological control landscape (Heimpel and Cock 2018). On one hand, we have a drastic drop in new introductions and therefore in the number of new classical biological control programs. On the other hand, we now have an incentive to search for native organisms that can be used in other methods of biological control (Cock et al. 2010). This process has important consequences for the relationships between biological control and taxonomy.
Classical biological control
With the widespread concern that biological control agents introduced outside their native range could negatively impact non-target populations, there has also been a shift in focus from classical biological control to other methods. Initial discussions on the relationships between taxonomy and biological control (e.g., Clausen 1942; Sabrosky 1955; DeBach 1960) took place at a time when classical biological control was the predominant approach, and less consideration was given to other methods. Currently, classical biological control faces numerous obstacles and has ceded its prominence to applied and, to a lesser extent, conservation biological control.
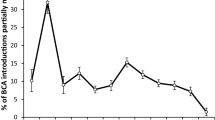
Two articles by Francis Howarth (1983; 1991) and the Convention on Biological Diversity (CBD 1992) are important milestones, as they reflect increasing awareness of the risks of classical biological control and the negative effects of invasive species on global biodiversity. Classical biological control is by definition an intentional introduction of an exotic agent that can establish in a new area and regulate populations of pest species in the long term (Eilenberg et al. 2001). The change in perception regarding the introduction of exotic species (Heimpel and Cock 2018) is associated with a drastic reduction in the number of new introductions from the 1980s onward, as demonstrated by Cock et al. (2016), as well as greater specificity of the agents introduced (Van Driesche and Hoddle 2016). In response to concerns raised, Food and Agriculture Organization (FAO) of the United Nations published the Code of Conduct for the Import and Release of Exotic Biological Control Agents (ISPM3) (FAO 1996), which led to significant advances in regulations, especially in developing countries (Kairo et al. 2003). More recently, the Nagoya Protocol on Access and Benefit Sharing (CBD 2011) also significantly affected the transit of organisms between countries (Cock et al. 2010). The restraints that surfaced after the signing of the Convention on Biological Diversity (CBD) impacted the exchange of natural enemies all over the world, as national regulations struggle to adopt the international standards for access to genetic resources and for export/import of live organisms (e.g., Coutinot et al. 2013).
As best practices for biodiversity protection, countries have adopted rules and regulations that favor selection of biocontrol agents that do not pose a risk to local biota, usually because they are highly prey/host specific or because they are not able to establish self-perpetuating populations, through implementation of regulatory procedures that include some kind of risk assessment (Barratt et al. 2010). Currently, risk assessments are mandatory in several countries, and should be mandatory in Brazil as well (Shimbori et al. 2022). Adaptation of regulations and legal restrictions for new imports is important to limit the impacts of new invasions. Hajek et al. (2016) commented that the lack of specific regulations has been a problem for microorganism biocontrol agents (viruses, bacteria, fungi, and nematodes), and highlighted the importance of the FAO ISPM3 guidelines (FAO 1996). In Brazil, since publication of the normative instruction IBAMA No. 5 (IBAMA 2016), importing exotic agents without already-established populations in this country requires a risk assessment by the national authority. Since the protocol for risk assessment is not yet established, currently there are no legal means to import a live organism for biological control. The road leading to an effective regulatory process is still long and winding for most nations, especially when harmonization among countries of economic blocs, for example, the Comitê Regional de Sanidad Vegetal del Cono Sur (COSAVE, South America) and the European and Mediterranean Plant Protection Organization (EPPO, Europe), is necessary to achieve this goal (Bale 2011). Geographically isolated countries and blocs made up of fewer countries, such as Australia and New Zealand and the North American Plant Protection Organization (NAPPO, North America), have more advanced regulatory processes (Hunt et al. 2008; Barratt et al. 2018a), in part because of the lower complexity involved.
With the exponential increase in international trade and the intensification of international travel, the resulting high propagule pressure tends to increase the rate of biological invasions (new introductions followed by colonization, expansion of the distribution area, and impacts on native populations) (Simberloff 2009). According to Pimentel et al. (2001), there are at least 120,000 invasive species of plants, animals, and microbes, which constitute one of the main threats to global biodiversity and are causing economic losses of over USD 300 billion annually, for only the six countries evaluated in that study (USA, UK, Australia, South Africa, India, and Brazil). The Invasive Species Specialist Group (ISSG 2018) concluded that the present context may stimulate countries to adapt their rules for importing natural enemies, so that the advantages afforded by classical biological control as a method of combating invasive species, whether by microorganisms or macroorganisms, can be used to the fullest. Lack of taxonomic knowledge of the biological control agents can delay the development of new biological control programs (Maddox et al. 1992; Zucchi 2002), a situation that may be aggravated by inadequate knowledge of the local biota and the low frequency with which pre- and post-introduction monitoring is conducted (Hoddle 2004; Barratt et al. 2006; Hajek et al. 2016).
Therefore, with the intensification of problems associated with biological invasions, an increasingly close collaboration among researchers in different areas of knowledge, i.e., taxonomy, biological control, and ecology, especially conservation ecology (e.g., invasion biology, niche modeling), regulators, and decision-makers will also be necessary. Experiences in earlier classical biological control programs that lacked stronger taxonomic studies, which resulted in failure or non-target impacts, have impacted the perception of these programs so that they are not seen as desirable options and are generally discouraged by regulations and agencies alike. However, international trade continues to intensify, with a consequent rise in introduction rates. As more invasive species are introduced, an increase in demand for classical biological control will follow, as the only viable solution for control in many cases, and this will not be limited to agricultural pests. The role of taxonomy and biological control in this scenario involving the import and release of natural enemies is crucial, due to the increased need for safer and sustainable solutions for the environmental, social, and economic risks imposed by biological invasions, including decreases in food supply, biodiversity, and health security (ISSG 2018).
Applied biological control
Applied biological control, especially inundation biological control, is growing rapidly, which may be associated with the necessity for replacement of the classical model, but also with the possibility of offering both sustainability and profitability at the same time. Even though this method was attempted as early as 1899 in Europe (Decaux 1899), and mass rearing was first used methodically in about the 1930s, in most regions, research on applied biological control as an alternative gained traction around the 1960s (DeBach and Hagen 1964; Ridgway and Vinson 1977; Parra 2014). This growth is often attributed to a greater general awareness of environmental issues following the publication of Silent Spring by Rachel Carson (1962) (e.g., Gay 2012).
In Brazil, the biological control market is growing at a fast pace. From 2019 to 2020, the market value of biological control products increased by 42% and from 2020 to 2021 is estimated to rise 33%, for a total net value of BRL 1.8 billion (CropLife Brasil 2022). These numbers are headed by microbial products but arthropods are also following this trend, and with that, the need is also increasing for taxonomic knowledge to identify species and to discover new biological control agents that can be incorporated into production systems. Thus, we have a scientific and technological interaction that proceeds from taxonomy to the market.
Certain characteristics are more relevant for the agents used in applied biological control than for those in classical biological control. Host specificity and establishment, for instance, are not considered necessary or even desirable. Although a few studies have sought to determine the impacts on non-target species, applied control tends to be considered safer, since it can use native species and establishment of self-perpetuating populations is not necessary, therefore having only transient impacts (Lynch et al. 2001; Kuske et al. 2003; Loomans 2021). However, Lynch et al. (2001) noted, based on a literature review, that effects on populations of non-target species are relatively frequent and that in 8% of cases they can be severe (i.e., reducing non-target populations by 40% or more). Therefore, before implementing this method, it is necessary to ensure that any impacts are in fact transient, by using either a non-establishing exotic agent or native agents.
Between 1980 and 2010, applied control developed into a highly organized industry, with at least 230 species available for commercial use; however, the total area treated is still very small, with enormous potential for growth (Collier and Van Steenwyk 2004; van Lenteren 2012; Parra 2014). Taxonomy is essential in helping to prospect for native biological control agents for mass rearing, avoiding the risks of unsuccessful imports, and feeding a rapidly growing industry. More attention to monitoring possible impacts is still needed and naturally must rely on collaborations between taxonomists and applied entomologists.
Regulating agencies also use taxonomic information for applied control. Licenses and registrations of biological agents for commercial use invariably depend on some confirmation of their taxonomic identity, and in many countries, applications are reviewed by experts, including taxonomists (e.g., Mason et al. 2017). In addition, voucher specimens are required to be deposited in permanent entomological collections to ensure that the identity of the registered organism can be reviewed in the future, should the need arise. In Brazil, the Joint Normative Instruction No. 1 of 2019 (MAPA/ANVISA/IBAMA 2019) requires specimens identified by taxonomists to be deposited in accredited collections in order to register an agent.
Another regulatory aspect of applied biological control is the need for quality control of products (natural enemies) produced and sold by companies, which must also be guaranteed by taxonomists. Taxonomic identifications are a necessary step in good-practice protocols for laboratories that produce biological control agents. In addition to consultations and reports by experts, rapid identification techniques can be very useful in certifying mass-reared insects, such as Loop-mediated isothermal amplification (LAMP) and Multiplex PCR, among others. These techniques have been used to identify agricultural pests, particularly groups that are difficult to identify accurately, such as Helicoverpa Hardwick (Lepidoptera, Noctuidae) (Amano and Nomura 2020), as well as for mass-reared natural enemies such as Trichogramma spp. (Dang et al. 2005; Davies et al. 2006; del Pino et al. 2013; Gonzales-Cabrera et al. 2014; Ivezić et al. 2018, 2021), and will be discussed in more detail in the section “Molecular tools for rapid identification.”
The relationship between applied biological control and taxonomy is as important as in classical control. This relationship provides an opportunity for collaboration between the two fields of research and a growing industry providing sustainable solutions for pest control. Two cases involving Trichogramma parasitic wasps illustrate the importance of this closer collaboration. First, Trichogramma galloi Zucchi has become one of the most common commercially produced species in Brazil, thanks to an exemplary model of close collaboration between taxonomy and biological control, as documented by Zucchi (2002). Second, in contrast, identification problems led to the unintended mass rearing and release of 300,000,000 individuals of Trichogramma fasciatum (Perkins) annually for 20 years to control Diatraea saccharalis (Fabricius) (sugarcane borer) in Barbados, because the populations had been mistakenly identified as Trichogramma minutum Riley (Gordh 1977).
Conservation biological control
The central priority of conservation biological control is to create a more complex and diversified agroecosystem conducive to beneficial interactions (Gliessman 2005). The ecological role of biodiversity in agroecosystems is the primary focus of research in this area (Altieri and Letourneau 1982; Andow 1991; Altieri 1999). Methods for conservation biological control are based mainly on culture techniques of habitat manipulation to improve control by the local fauna of natural enemies or by inducing different responses of herbivores to plant diversity (Gurr et al. 2004; Fontes et al. 2020). Conservation biological control is considered an excellent alternative to foster more sustainable agroecosystems, as it requires lower levels of input and promotes higher diversity. However, as this is a complex strategy operating on multiple scales, many issues affecting suppression by natural enemies must be considered and possible limiting factors identified for effective application (Gillespie et al. 2016; Begg et al. 2017). The higher diversity in these agroecosystems requires greater taxonomic knowledge, which may be a limiting factor per se.
As social demands on sustainable production increase (Tracy 2014; Altieri and Nicholls 2020) and options for classical biological control have become restricted, conservation biological control tends to play a larger role in agroecosystems, relying on collaborative research in ecology (including natural enemy dispersal, habitat connectivity and other landscape effects, and trophic interactions), chemical ecology, genetics, insect behavior, and socioeconomic aspects, among others. This multidisciplinary area, encompassing such a vast array of disciplines, is necessarily based on taxonomy, which is essential for recognition of the species of natural enemies and insect pests present in the systems. A vast literature has addressed this ecological service through conservation biological control (for example, Gliessman 2005; Venzon et al. 2021), which has taxonomy as a basic foundation for increasing functional biodiversity.
Current status of the biological control–taxonomy relationship
In general, laws regulating the import and release of biological control agents were tightened from 1980 to 1990. With the possible exception of New Zealand and developing countries where the use of biological control is more recent and the adoption of FAO’s ISPM 3 has facilitated regulation (Kairo et al. 2003), it has become much more difficult to introduce exotic species worldwide, including for biological control purposes. Even countries with advanced legislation require lengthy import control processes. Although the number of new introductions has decreased drastically since the 1990s, an increase may become possible after scientific advances in the area, in response to current demand. For example, considering applied biological control in this situation, a company could be discouraged from importing a species with a ready (foreign) technological package, due to the many regulatory requirements. An illustrative example, in this case with a pollinator, is the import and release of the bumblebee Bombus terrestris (L.) (Hymenoptera, Apidae), native to Eurasia and Northern Africa, into South America (Smith-Ramírez et al. 2018; Fontúrbel et al. 2021). After being repeatedly introduced in Chile, it has invaded Argentina and may reach Brazil and Uruguay (Acosta et al. 2016), even though its import was rejected in these countries (Velthuis & van Doorn 2006). This demonstrates the importance of laws that regulate the entry of exotic species (Aizen et al. 2019) and the consequent incentive to prospect for native species, as exemplified by a research grant for bioprospecting Brazilian species of Bombus Latreille for pollination in agriculture, awarded in 2018 by the State of São Paulo Research Foundation (FAPESP) (FAPESP no. 16/21555–9; available at https://bv.fapesp.br/pt/auxilios/100336/abelhas-bombus-para-programas-de-agricultural-pollination-bioprospection-of-native-species-of-Brazil/).
Importations of non-native Trichogramma species furnish other examples, because studies on the local fauna of many regions are still incipient, even though this knowledge would be important for furthering the use of Trichogramma in biological control programs (Querino et al. 2010). In almost all habitats, native species adapted to local conditions are already present, an indication that natural parasitism by Trichogramma could be high (Pinto 1999). Cock et al. (2010) confirmed a trend toward an increase in the use of native species. Based on the European market, the authors found that the proportion of native species used for augmentative control grew from 42% in 1960–1989 to 76% in 2000–2009. These trends in the European market may be followed in other parts of the world as regulations became increasingly restrictive, encouraging improvement of taxonomic knowledge and adaptations from the applied biological control industry.
An excellent recent example involving invasive pests, exotic biological control agents, and regulations illustrates how the current situation increases the importance of closer collaboration between applied entomologists and taxonomists. The Asian brown marmorated stink bug, Halyomorpha halys (Stål) (Hemiptera, Pentatomidae), became a serious pest after it became established in Europe and the USA. A classical biological program was initiated in 2005 in the USA by Kim Hoelmer and collaborators, who found an excellent candidate biocontrol agent in Asia, the samurai wasp, Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) (Conti et al. 2021). Years of research in a quarantined facility apparently did not generate data indicating that the samurai wasp could be released in the USA without risks to the environment, which is estimated in terms of non-target effects (Louda et al. 2003). However, in 2014, T. japonicus was found in Maryland (Talamas et al. 2015), as a result of Hoelmer’s collaboration with a team searching for native parasitoids on stinkbug eggs (Buffington et al. 2018). This serves as a warning that natural enemies might be inadvertently introduced along with their hosts and of the need for regulations that allow timely screening and assessment of safe and efficient agents, especially considering the increasing flow of international trade. This also shows that close collaboration between a taxonomist and applied entomologists was essential while searching for native agents for biological control. Moreover, this illustrates the role of government agencies and national authorities in plant protection, providing the structure and organization necessary for this relationship to flourish.
The above examples demonstrate the incentive for taxonomy related to native natural enemies. We believe this is a transforming force to strengthen this research field and its relationship with biological control. However, considering the “unnaturally” wide distribution range of many pests and other anthropophilic species, more attention is needed in situations where exotic species are already adapted to a new environment. As border control and quarantine lists are prepared to detect invasive species, natural enemies may also become invasive and identification of species originating from another part of the world may be difficult for many reasons, mostly related to the lack of taxonomic studies. Molecular markers may be the best tools for aiding taxonomy and biological control in such cases.
In conclusion, the legal restrictions intended to protect biodiversity that hinder or prevent importation of species obviously affect efforts for classical biological control, but also impact applied biological control, since the search for solutions would be more and more pushed toward native (or exotic established) species, as the risk assessment and monitoring procedures for introduced species, when they exist, are lengthy and costly. However, for taxonomy, this seems to be a very favorable scenario for further prospecting work that will necessarily involve greater knowledge of the local fauna, and therefore a greater demand for taxonomists. Similarly, regarding the examples of Bombus terrestris and Trissolcus japonicus, when prospecting for native agents was encouraged, further incentives in this direction should be expected. This is a very positive situation for conservation biological control, as it does not use exotic species and depends on knowledge of native species, which will be encouraged.
Molecular taxonomy and databases
Molecular markers and their public databases are currently considered highly useful tools in several areas of knowledge. Much of this usefulness relies upon correct association between names and genetic sequences (Schoch et al. 2020). As Strasser (2008) commented, public databases and the use of molecular markers in taxonomy have led to a convergence of natural history practices and the experimental sciences. DNA barcoding, the most prominent of molecular markers used in studies of insects, was proposed originally with the aim of becoming a universal tool for identifying eukaryotic organisms, at the species level, from a DNA sequence (Hebert et al. 2003a). For identification, barcoding would depend on the construction of a database to store sequences of all described species, as well as species not yet described, called the Barcode of Life Database Systems (BOLD Systems) (Ratnasingham and Hebert 2007). This and other molecular markers, and more recently genome-scale studies based on Next Generation Sequencing (NGS), have revolutionized taxonomy, in particular allowing highly refined species delimitation and reconstruction of phylogenies with maximum support, based on a previously unimaginable amount of information. This revolution obviously has profound consequences for collaborations among taxonomists, molecular biologists, and applied entomologists.
S.E. Miller (2007) called the advent of the DNA barcode a taxonomy renaissance. In the area of applied entomology, barcoding could make identification faster and more accessible to anyone, even in the field. To make available such a free tool, for example, via an internet-content search engine such as Google, would require a large investment in developers and means of generating profits that would be reinvested in further development. The main “developers” of these databases are taxonomists who curate information by associating names with DNA sequences based on examination of voucher specimens, and then generating reference sequences that are the basis of DNA barcoding. However, and as is recognized as a bottleneck in barcoding logistics (Borisenko et al. 2009), taxonomists do not receive due recognition for their work or, usually, any kind of compensation. It would be useful for the investment strategies in what S.E. Miller (2007) called the “Taxonomic Enterprise” to be restructured, in order to value and promote taxonomic curation so that all the benefits of public databases could be fully achieved.
Despite the herculean, and essentially impossible, task of sequencing all living species, scientists are increasingly using and expanding the data available for these markers, and the main databases are growing exponentially. DNA barcoding alone (COI) reached 2.5 million sequences in 2018 in the GenBank database (Porter and Hajibabaei 2018), while BOLD reached 11 million COI sequences in 2022, representing approximately 385,000 described species (BOLD 2022) and constituting an invaluable information resource for science. These numbers, although impressive, show that most species are still not represented. The lack of reference sequences is a major problem that is even worse for many poorly known taxa, even at high taxonomic levels. Initiatives to mitigate this problem have emerged, with the potential to use museum specimens (Santos et al. 2022).
An important difference between the BOLD and GenBank databases, which are the two main public databases for DNA barcoding, is that BOLD can be considered a sequence curation tool while GenBank is a repository (Meiklejohn et al. 2019). The issue is recognized by the community, and efforts to improve annotation errors have been proposed for GenBank, but the quality still depends on accuracy and updates from submitters (Bidartondo et al. 2008; Schoch et al. 2020). The few studies reviewing the reliability of the databases regarding taxonomic identification have described some worrying trends. For example, studies on fungi found that 80% of the sequences lack reference vouchers (Bridge et al. 2003; Nilsson et al. 2006). Even with the stricter requirements of BOLD, Meiklejohn et al. (2019) calculated a relatively low accuracy for insect identification (~ 35%), lower than the GenBank BLAST tool (53%). Thus, although there is a premise that the deposited sequences should be obtained from voucher specimens identified by an expert, the chance of error when using these databases cannot be ignored, as they are public databases. The lack of association between sequences and taxonomic names, whether due to misidentification, lack of comparison with morphologically described species, or simply not assigning a name, is a problem that needs to be addressed and systematized in order to achieve reliability in the use of databases. The debate on this and other points relative to the use of molecular markers in taxonomy is quite intense and extensive (e.g., Meyer and Paulay 2005; Cameron et al. 2006; Begerow et al. 2010; Zhang et al. 2012). The present discussion serves merely to note that end users, applied entomologists especially, should remain aware of these faults, without discouraging the use of markers, as the advantages of this genetic information within the scope of biological control outweigh any of the shortcomings and may not be directly affected by them if the data are treated with due care.
Examples of identification problems in databases are very common—what Collins and Cruickshank (2013) call the “second sin” of DNA barcoding—and can slow the progress of biological control research in very harmful ways. In a recent survey, we found that the name Trichogramma pretiosum Riley, which corresponds to a well-studied species for applied biological control of several of lepidopteran pests, is attributed to 6 distinct molecular taxonomic units (MOTUs) according to the Barcode Index Number (BIN), which could be equivalent to species, for only 75 deposited barcodes. For Telenomus podisi Ashmead (Hymenoptera, Scelionidae), a parasitoid that has been intensively studied for the control of pentatomids, the number of BINs reaches 13 for the ~ 3600 barcodes deposited. Therefore, an unambiguous identification based on DNA barcoding is currently impossible, as is determining which of these MOTUs corresponds to the original names Tr. pretiosum and Te. podisi. It is likely that control programs are being conducted with incorrectly identified wasps, or even that other species are being identified under the names Tr. pretiosum and Te. podisi. Again, the question “What difference does it make, if control is reached?” is pertinent. Among the numerous problems, we can mention the erroneous accumulation of information about biology, ecology, and distribution linked to a species, when this information actually refers to other species. This can lead to a false impression of polyphagy and consequent interruption of classical biological control programs, or even to the attempt to use an agent in applied control of a pest outside the host range of the agent. To put it into perspective, this is exactly the type of problem described by researchers in the past (e.g., Clarke 1993). Howarth (2000) cited the example of sciomyzid flies introduced into Hawai’i for snail control. The program, aiming to introduce the marsh fly Sepedon sauteri Hendel (Diptera, Sciomyzidae), ended up introducing 11 species of flies, and reexamination of the vouchers showed that none of them seemed to be S. sauteri (Hardy and Delfinado 1980). Because a permit was issued for that species, there would be no problem with future introductions of S. sauteri, even if the actual species has not established. Well-curated reference sequences would prevent these problems.
Besides identification, DNA barcoding has also been used in taxonomy as a tool for species delimitation and is regarded as a fundamental link between traditional taxonomy and the “DNA world” (Hubert and Hanner 2015). However, contrariwise, there is a lack of connection between the “traditional” taxonomy and the “molecular” taxonomy along the lines proposed by Sharkey et al. (2021), that is, replacing the entire description with only a genetic sequence, a couple of photographs and a list of material examined, which seems worrying to us. Generating and analyzing sequences that are considered unique and assigning names to these sequences could drastically accelerate knowledge of biodiversity (Godfray 2002; Hebert et al. 2003b). However, applying the methods used by Sharkey et al. (2021) would generate this knowledge at the expense of loss of centuries of scientific history, as the names and all scientific research linked to these names cannot be accessed for species without available molecular markers. Unfortunately, this is still the case for the vast majority of species already described (see, however, Santos et al. 2022). Meier et al. (2022) discussed the issues of the approach taken by Sharkey et al. (2021), pointing to the increasing availability of large throughput imaging and sequencing, and the advances in algorithms that can integrate multiple sources of data, as sound solutions for the taxonomic impediment, by providing taxonomists with a solid foundation for species descriptions.
There are serious criticisms of the way that taxonomic units are delimited based on molecular markers, and whether there is coherence between these methods and species concepts, so that species can be explicitly delimited by comparison of their genetic sequences (e.g., DeSalle et al. 2005; Meier et al. 2006; Rubinoff et al. 2006; Lowenstein et al. 2009; Zamani et al. 2021). Therefore, combination of different models with transparency and reproducibility of the methods is strongly recommended (Meier et al. 2022). Additionally, a combination of multiple loci may be necessary for delimitation (Heraty 2004, 2009). Some examples from parasitoids, such as the Peristenus pallipes complex (Zhang et al. 2012), the Aphelinus varipes complex (Heraty et al. 2007), and certain species of Trichogramma (Stouthamer et al. 1999; Pinto et al. 2002, 2003; Cruaud et al. 2018, 2019), demonstrate that divergence in a single gene region may not correspond to reproductive isolation, and even the use of more than one region may not be sufficient (e.g., Zhang et al. 2017), requiring further mating-compatibility studies in different populations (e.g., Vickerman et al. 2004). These problems are inherent to the field of taxonomy, in regard to the evolutionary process and its multiple levels, and demonstrate that in many cases there is no simple solution other than a closer collaboration among scientists in several fields, including taxonomy, ecology, evolutionary biology, applied entomology, and even behavioral ecology.
Criticisms aside, it would be extremely advantageous, not only from the point of view of biological control, if the taxonomic challenges posed by all pests and biological control agents could be solved by obtaining a DNA sequence. The entire movement around the development of these markers, such as DNA barcoding, revolves around this possibility. The identity of an organism, however, depends on the taxonomic work involved in attaching the name to the marker. Without this work, the only possible statement is that the sequence obtained is equal to some, or different from all, of the sequences deposited in the databases. Therefore, it makes no sense to name sequences, and so taxonomic work remains relevant in all its complexity. This is a long-standing discussion (e.g., Tautz et al. 2003; Lipscomb et al. 2003; Wheeler and Valdecasas 2005; Hickerson et al. 2006; Carvalho et al. 2007; Packer et al. 2009; Zamani et al. 2021) that is beyond the scope of the present article, although some points are still controversial. However, it is necessary to point out that solutions such as DNA taxonomy or cybertaxonomy are proposed to accelerate taxonomy but depend on the growth of taxonomic foundation. Therefore, we agree with Carvalho et al. (2005) and Engel et al. (2021), in that fighting taxonomic impediment instead of creating shortcuts for the lack of taxonomists is the only way to achieve meaningful goals for any science based on biodiversity.
In the context of biological control, we can imagine a scenario that illustrates the possibilities and limitations of using molecular markers and databases, as follows:
Imagine a Brazilian research group that finds a natural enemy with high potential for use in biological control. The species cannot be identified immediately, so the first step is to obtain sequences from a molecular marker, and here the DNA barcode is chosen. The sequence is compared to the sequences in BOLD and GenBank and an identical sequence is found. The researchers, here in Brazil, discover that the other sequence was deposited by researchers in South Africa. Both groups are studying the same pest, which has a worldwide distribution, and the parasitoid apparently dispersed along with the pest. The two groups begin to exchange information and develop an international research program for mass rearing and release of this natural enemy. Groups in Europe and Australia are are evaluating the possibility of importing specimens for introduction. Finally, the only active taxonomist specializing in the taxon responds to the request to identify the parasitoid. While in the process of retiring, he receives the material and responds with a name and a publication attached: "This species, Aa aa, described by me 40 years ago, is present in all biogeographic regions (except Antarctica) and was introduced from Europe to the Americas, Africa, and Australia. It was studied for decades without success for the control of the target pest. In the 1990s the studies were abandoned because its release did not result in a significant reduction of the pest populations." Most programs did not keep specimens and the only voucher specimens from these research programs are in very poor condition and scattered in collections around the world, except for material studied by the taxonomist. In conclusion, applied entomologists should use protocols similar to those followed by insect collectors, that is, collecting, assembling, tagging, and preserving specimens, which will be of great value in any biological control programs that may be implemented at any time.
An inherent advantage in comparing molecular markers is their universality and accessibility. The process of detecting that the same entity occurs at isolated points on the planet is fast and reliable. In the above hypothetical example, this could immediately draw researchers’ attention to the disjunct distribution, possibly reflecting human action. This feature is very useful, especially for species that live in proximity to or associated with anthropic environments, such as pests and their natural enemies (Andersen et al. 2019).
If, in this hypothetical example, the natural enemy species had been given a second name, Aa bb, in a taxonomic study based exclusively on molecular markers, the problem would not have been noticed. The species would be identified as Aa bb by the association between the marker and the name; however, all previous knowledge of the species Aa aa would be ignored and there would even be a risk of attributing the origin of the species to a place where it was introduced. Even if it were technically possible to mass-rear and release such a species with modern techniques, the valuable knowledge already generated would have been wasted. Likewise, if the sequences in databases had been incorrectly assigned to a different species, all sorts of problems related to misidentification could follow. As DeBach (1960) commented, “a natural enemy species, perhaps undescribed and new to science, but wrongly given the name of a described species, may be as effectively hidden as if it was never collected.”
NGS and museomics
The increasing ease of obtaining genome-scale data following rapid advances in High Throughput Sequencing technology is generating new and exciting possibilities (e.g., Wen et al. 2017). Some have claimed that this new genomic era could mean the end of DNA barcoding (Taylor and Harris 2012; Joly et al. 2014), while others have highlighted the complementarity of genomics and DNA barcoding (Grant et al. 2021). In fact, most of the applications of genomics in taxonomy are related to the reconstruction of phylogenies, improving our understanding of the evolution of life and consequently our classifications. This knowledge can be particularly important in risk assessments, as discussed by Barratt et al. (2018b). It is possible, however, for an extended concept of DNA barcoding to integrate genomic data (Coissac et al. 2016). For example, Allio et al. (2020) developed a pipeline for mitogenomic data extraction from UCE (Ultra Conserved Elements) libraries, retrieving COI barcoding and confirming identifications with 100% success. As demonstrated by Cruaud et al. (2019), it is now possible to generate genomic data from minute parasitoids, which can be important for molecular taxonomy in support of biological control.
The emerging field of historical DNA or museomics is being supported by NGS techniques (Raxworthy and Smith 2021), for example, Target Enrichment using UCE, because of the high rates of success for degraded samples (Blaimer et al. 2016; Zhang et al. 2019). Mini-barcodes have also proved useful for museum specimens (Van Houdt et al. 2010). Research in this area will be essential to fill the gaps in reference sequences in DNA databases for described species, improving both species identification and discovery.
Molecular tools for rapid identification
Numerous diagnostic methods based on molecular-biology techniques have been developed in the last two decades. Some have recently become popular because of their usefulness in diagnosing COVID-19 (Amaral et al. 2021), such as LAMP (Loop-mediated isothermal amplification), developed for rapid amplification of DNA with high specificity using six different primers (Notomi et al. 2000). LAMP is currently considered one of the most versatile methods for identifying organisms, as it does not require equipment for electrophoresis or real-time monitoring. This method is accurate, inexpensive, and can be used for on-site identification, such as during monitoring surveys (Notomi et al. 2015). Other examples are PCR–RFLP and multiplex PCR, which can distinguish among numerous target species with conventional PCR equipment but are not adapted for large samples. On the other hand, real-time PCR and ddPCR are excellent alternatives for rapid analysis of multiple samples but require expensive equipment (Amano and Nomura 2020).
These methods are increasingly used to identify pests and invasive species. LAMP, for example, is being used to identify quarantined species on-site (von Felten 2017, in Switzerland), especially fruit flies (Blaser et al. 2018; Sabahi et al. 2018; Blacket et al. 2020; Kitano and Takakura 2020; Dermauw et al. 2022), using a portable device. LAMP is also helpful for field identification, especially when specimens cannot be reliably identified using traditional methods (e.g., a stage or sex is not morphologically useful for accurate identification): Agarwal et al. (2020; 2022) for the fall armyworm, Spodoptera frugiperda (J.E. Smith), and the grape phylloxera, Daktulosphaira vitifoliae (Fitch); Rako et al. (2021) for the Khapra beetle, Trogoderma granarium Everts. All these methods only detect predefined targets for which the primers need to be designed, and may not be sensitive for some lineages such as biotypes (e.g., Hsieh et al. 2012). Therefore, development of tools for biological control, and more broadly for integrated pest management (IPM), requires close collaboration between taxonomists and molecular biologists.
The same technology can be used to detect infections by Wolbachia (Gonçalves et al. 2014) and other symbiont bacteria that might reduce the ability for establishment and biological control due to cytoplasmic incompatibility (Sinkins et al. 1995). The ability to detect infections is quite useful, considering new introductions and the effect of bacterial symbionts on the life history of their hosts, as exemplified by the introduction of Encarsia spp. into the USA (Gebiola et al. 2016b). This example involves a complex taxonomic history of two cryptic species, Encarsia inaron (Walker) and E. partenopea Masi, both introduced into the USA as populations from two different localities, but under the same name. Gebiola et al. (2016b) demonstrated not only that these are two species, but also that they are infected by different bacteria, causing cytoplasmic incompatibility, one by Wolbachia and the other by Cardinium. This case draws attention to the risks of mixing populations with different bacteria that may cause reproductive failure, or any important change in fitness, in imported biological control agents. Luckily, the populations imported into the USA were from two different, reproductively isolated species, and that particular biological control program is considered a textbook example of success.
Rapid molecular diagnostics is useful for quality control in biofactories. For parasitoids, most published studies have used Multiplex PCR for rapid identification of Trichogramma species (Dang et al. 2005; Davies et al. 2006; del Pino et al. 2013; Gonzales-Cabrera et al. 2014; Ivezić et al. 2018, 2021), with some exceptions in Braconidae (Gariepy et al. 2005) and Encyrtidae (Rugman-Jones et al. 2011). Studies with LAMP are more recent, and this technique is more widely used in pest detection (e.g., Jenkins et al. 2012), with some examples for parasitoids such as the study by Nam et al. (2022) to identify two species of Diadegma Foerster (Ichneumonidae) that parasitize the diamondback moth, Plutella xylostella.
Recent examples
Numerous control programs have demonstrated the importance of population-level studies in the evaluation of agents. This has been discussed since the first papers on the relationship between taxonomy and biological control (e.g., Sabrosky 1955). A classic example of the importance of considering strains and populations in biological control occurred in New Zealand, when the beetle parasitoid Microctonus aethiopoides Loan (Hymenoptera, Braconidae) was imported and released at two different times, to control two different pests, and achieved different outcomes, demonstrating the importance of taxonomy in risk assessment for biological control. The first strain has a broader host range and had success in controlling the lucerne weevil, Sitona discoideus Gyllenhal (Coleoptera, Curculionidae) (Goldson et al. 1990; Ferguson et al. 1994; Barratt et al. 2018b), but commonly attacks native species (Aeschlimann 1995; Barratt et al. 1997). The second strain was imported after careful risk assessment, has a narrow host range, and achieved relative success in controlling the clover root weevil, Sitona obsoletus (Gmelin), with no significant non-target effects (Goldson et al. 2005; Gerard et al. 2011; Basse et al. 2015). Since the early studies, infraspecific variation within M. aethiopoides has received some attention (e.g., Aeschlimann 1983a, 1983b; Adler and Kim 1985), although not primarily concerned with host range and non-target effects (Aeschlimann 1980). The difference in outcome related to non-target effects can be attributed to considerably better knowledge of the parasitoids’ infraspecific taxonomy, the phylogeny of its weevil hosts, the host preferences of each parasitoid strain (Phillips et al. 2008; Barratt et al. 2018b), and better regulation processes.
However, in taxonomic practice, the importance of the species as a fundamental unit predominates. Even if the species concept is well established, it may be insufficient to identify populations with desirable characteristics for biological control. A more in-depth study may be needed to reveal cryptic species, biotypes, or populations with relevant adaptations. Therefore, integrative taxonomy that addresses intraspecific variation, using phylogeographic analyses, for example, can be decisive to achieve satisfactory results in a close collaboration between taxonomy and biological control (Huang 2020). A series of studies on Trichogramma has demonstrated the importance of data integration, not only for species delimitation and reproductive isolation within a species complex (e.g., Pinto et al. 2003), but also for demonstrating intraspecific variability in certain parasitoids, such as Trichogramma marandobai Brun, Moraes and Soares (Vieira et al. 2015) and Tr. pretiosum (Querino et al. 2002; Viana et al. 2021).
A study on euphorines (Hymenoptera, Braconidae) exemplified this situation using integrative taxonomy, where molecular data predominated in species delimitation (Zhang et al. 2017). The authors concluded that a complex of nine nominal species of the Peristenus pallipes group, previously delimited essentially based on ecology and morphology (Goulet and Mason 2006), was in fact composed of only three species. This conclusion has important consequences for biological control, as well as for the definition of conservation priorities, since the species as previously understood would have more-restricted distributions and host ranges. Zhang Y.M. et al. (2017) recognized the importance of studying the group at a population level, investigating microevolution within the complex. However, with current knowledge, it is impossible to determine whether at least some of the previously recognized species actually correspond to distinct populations or lineages. Likewise, although a complex of cryptic species that are notoriously difficult to distinguish were more clearly delimited, an even greater level of detail would be necessary to select biological control agents. We agree with Huber et al. (2021) that the classification proposed by Goulet and Mason (2006) may be more useful to select strains for biological control.
The taxonomic revision of Trichopoda Berthold (Diptera, Tachinidae) by Dios and Nihei (2020) reveals the complexity of this topic, as they showed that a group of species widely studied in biological control is a polymorphic species (quite the opposite of a group of cryptic species), while differentiating other species of this group, previously considered cryptic, based on morphology. The genus Trichopoda has a native distribution in the Nearctic and Neotropical regions. This recent revision demonstrated the extremely complex taxonomic history of the group, with 16 synonyms for Trichopoda pennipes (Fabricius), including names also used in biological control, such as T. giacomellii (Blanchard) (Dios and Nihei 2020). Trichopoda pennipes was supposedly introduced into several countries to control Nezara viridula (L.) (Hemiptera, Pentatomidae), a pest with worldwide distribution and important in numerous crops, especially soybean. Instead of the widely distributed and well-studied T. pennipes, the species that actually established populations in Europe was Trichopoda pictipennis Bigot (Dios et al. 2021). The entire history involving these species began a century ago, due to the interest in controlling a serious agricultural pest, and at that time the identity of accidentally introduced organisms had not been established. Notwithstanding, modern taxonomic studies have not been able to elucidate the nature of the populations and possible subspecies that compose T. pennipes, so that the differences among these lineages, which may be important for biological control, are still obscure. This example demonstrates the lack of studies on important and taxonomically challenging groups, as well as the limitation of taxonomic studies based only on morphology, without molecular and ecological data (Packer et al. 2009). Complex groups with cryptic and polymorphic species, and important differences at the infraspecific level, as in Trichopoda, should be studied using a combination of methods to clarify the extent of population and species diversity.
In the last two examples, the lack of close collaboration with applied entomologists may have rendered taxonomists less sensitive to the level of refinement required in biological control, which is often on the threshold between cryptic species and populations (subspecies) or geographic forms within a species with distinct biological characteristics such as preferences in oviposition and host range (e.g., Pinto et al. 2003; Goldson et al. 2005; Huang 2020; Seehausen et al. 2020). In view of the enormous gap in knowledge of species, which is considerably larger for parasitoids and microorganisms, and the taxonomic impediment, particularly the lack of professionals with training in taxonomy, taxonomists may find themselves faced with the need to decide between dedicating their time to detailing nuances of a species’ microevolution or to revising a larger group such as a genus or tribe. Choosing the second option seems naturally more acceptable at this juncture. However, in some cases, taxonomists have been able to solve questions at the three taxonomic levels (alpha, beta, and gamma), largely using molecular data, but also based on morphological knowledge and other sources. The Euphorinae (Braconidae) provide some good examples, where the identities of species of Peristenus Foerster (Zhang et al. 2017) and Eadya Huddleston and Short (Peixoto et al. 2018; Ridenbaugh et al. 2018), two important genera in biological control, were clarified concurrently with the phylogeny of the subfamily (Stigenberg et al. 2015; Zhang et al. 2018).
An essential practice: taxonomic validation
Taxonomic validation, as defined by Packer et al. (2018), is a three-step process during the identification of an organism in any study using scientific names, in order to confer falsifiability on identifications presented in publications. In general terms, taxonomic validation consists of (i) a description of the identification procedure, (ii) an explanation of the concept used for a given taxon, and (iii) a designation of a collection for deposit of voucher material. As most studies based on biodiversity, including applied studies, have species as their fundamental unit, the possibility of replicating and falsifying identification results through correct and explicit methods must be an integral part of sound scientific activity. However, only 1.8% of studies in entomological journals evaluated by Packer et al. (2018), on various subjects, reported conducting these three steps that allow replicability of the taxonomic information. This is alarming, since the possibility of changes in taxonomic concepts, together with potential identification errors, can make a significant part of published information completely unverifiable and thus unusable. With taxonomic validation, it is possible to trace back and correct errors without losing information, either by simply verifying that the concept used is outdated or by checking voucher specimens.
Deposit of molecular marker sequences in public databases is another very useful practice in the taxonomic validation process, and should be followed whenever possible, along with citation of accession numbers in publications. Importantly, even in cases where the identification has been carried out by means of comparison with molecular markers, it is essential that all three steps be taken. In this case, the species concept must be explained upon citation of taxonomic works that associate the sequences of markers with the names, given the lack of reliability of public databases.
In order to clarify the species concept used, and in some cases as part of the description of the identification procedure, it is necessary to cite taxonomic works that enable identification of taxa whose names are mentioned in the publication. The suggestion by Packer et al. (2018) is quite timely, as it allows tracing the taxonomic concept underlying the identification, based on the most relevant bibliography at the time, that is, not necessarily the original description, but the work that contains the most recent concept followed by the authors. This is expected to alleviate one of the primary aspects of the crisis in taxonomy, through the considerable increase in the number of citations.
Numerous articles have discussed the problem of lack of citations of taxonomic articles (e.g., Drew 2011; Meier 2017), despite the undeniable importance of this field (Mayr 1968; Danks 1988). Here, we must emphasize that the common practice of not citing taxonomic articles is not only unjustified but also harmful to taxonomy and to the proper development of biological sciences that have biodiversity as their primary focus of study. The discovery, delimitation, and naming of a species, as well as the creation of tools for its identification, open a multitude of possibilities of use for this name and all the scientific communication surrounding it. Imagine if Trichogramma galloi, currently one of the most commercially important species in Brazil for biological control, had not been studied and described in such a way as to make its identity clear, what would have become of the over two thousand scientific articles on this species? Adding the original description (Zucchi 1988) and the main published identification keys (Querino and Zucchi 2005, 2011, 2019), these taxonomic works have been cited 93 times,Footnote 2 which means that less than 5% of the publications using the name Tr. galloi cited references that could help to ensure the correct use of the name as part of their results.
We understand taxonomic validation to be essential for both taxonomy and biological control, and more broadly for the biological sciences. All aspects of this process are customary practices in taxonomy, which denotes the advantage of close collaboration between applied entomologists and taxonomists and tends to improve the taxonomic validation process in publications in applied entomology, which is often overlooked. Much has been said in recent years about a taxonomic renaissance (e.g., S.E. Miller 2007), or even a new era of cybertaxonomy (e.g., Wheeler 2007; Hita-Garcia et al. 2019). However, the problems related to the “taxonomic impediment” remain serious and the crisis in taxonomy is still worrying. Fortunately, many journals have begun to require one or more steps of taxonomic validation in their publications, which we regard as one of the main actions toward valuing taxonomy by peers. These steps should increase the number of citations and show the true importance of taxonomic research as an enabler of rather than an impediment to work focused on biological diversity.
Data Availability
All data generated or analysed during this study are included in this published article.
Notes
-
The term “augmentation” biological control is widely used in the literature, mostly referring to inundation biological control, as defined by Eilenberg et al. (2001). Here, we use the term “applied” biological control to refer to both inoculation and inundation biological control, even though most of the examples used in applied biological control are for inundation.
-
Based on a search in Google Scholar, accessed on June 13, 2022.
References
Acosta AL, Giannini TC, Imperatriz-Fonseca VL, Saraiva AM (2016) Worldwide alien invasion: a methodological approach to forecast the potential spread of a highly invasive pollinator. PLoS ONE 11(2):e0148295. https://doi.org/10.1371/journal.pone.0148295
Adler PH, Kim KC (1985) Morphological and morphometric analyses of European and Moroccan biotypes of Microctonus aethiopoides (Hymenoptera: Braconidae). Ann Entomol Soc Am 78:279–283. https://doi.org/10.1093/aesa/78.3.279
Aeschlimann J-P (1980) The Sitona (Col.: Curculionidae) species occurring on Medicago and their natural enemies in the Mediterranean region. Entomophaga 25:139–153. https://doi.org/10.1007/BF02374316
Aeschlimann J-P (1983a) Notes on the variability of Microctonus aethiopoides Loan (Hymenoptera: Braconidae; Euphorinae). Contrib Am Entomol Inst 20:329–335
Aeschlimann J-P (1983b) Sources of importation, establishment and spread in Australia, of Microctonus aethiopoides Loan (Hymenoptera: Braconidae), a parasitoid of Sitona discoideus Gyllenhal (Coleoptera: Curculionidae). J Aust Entomol Soc 22:325–331. https://doi.org/10.1111/j.1440-6055.1983.tb02111.x
Aeschlimann J-P (1995) Lessons from post-release investigations in classical biological control: the case of Microctonus aethiopoides Loan (Hym., Braconidae) introduced into Australia and New Zealand for the biological control of Sitona discoideus Gyllenhal (Col., Curculionidae). In: Hokkanen HMT, Lynch JM (eds) Biological control: benefits and risks. Cambridge University Press, New York, pp 75–83
Agarwal A, Cunningham JP, Valenzuela I, Blacket MJ (2020) A diagnostic LAMP assay for the destructive grapevine insect pest, phylloxera (Daktulosphaira vitifoliae). Sci Rep 10:21229. https://doi.org/10.1038/s41598-020-77928-9
Agarwal A, Rako L, Schutze MK, Starkie ML, Tay WT, Rodoni BC, Blacket MJ (2022) A diagnostic LAMP assay for rapid identification of an invasive plant pest, fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae). Sci Rep 12:1116. https://doi.org/10.1038/s41598-021-04496-x
Aizen MA, Smith-Ramírez C, Morales CL, Vieli L, Sáez A, Barahona-Segovia RM, Arbetman MP, Montalva J, Garibaldi LA, Inouye DW, Harder LD (2019) Coordinated species importation policies are needed to reduce serious invasions globally: the case of alien bumblebees in South America. J Appl Ecol 56(1):100–106
Allio R, Schomaker-Bastos A, Romiguier J, Prosdocimi F, Nabholz B, Delsuc F (2020) MitoFinder: efficient automated large-scale extraction of mitogenomic data in target enrichment phylogenomics. Mol Ecol Resour 20:892–905. https://doi.org/10.1111/1755-0998.13160
Altieri MA (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74:19–31. https://doi.org/10.1016/B978-0-444-50019-9.50005-4
Altieri MA, Letourneau DK (1982) Vegetation management and biological control in agroecosystems. Crop Prot 1(4):405–430. https://doi.org/10.1016/0261-2194(82)90023-0
Altieri MA, Nicholls CI (2020) Agroecology: challenges and opportunities for farming in the Anthropocene. Cienc Investig Agrar 47(3):204–215. https://doi.org/10.7764/ijanr.v47i3.2281
Amano T, Nomura M (2020) A diagnostic loop-mediated isothermal amplification method to distinguish Helicoverpa armigera (Lepidoptera: Noctuidae) from other related species in the New World. J Insect Sci 20(10):1–6. https://doi.org/10.1093/jisesa/ieaa046
Amaral C, Antunes W, Moe E, Duarte AG, Lima LM, Santos C, Gomes IL, Afonso GS, Vieira R, Teles HSS et al (2021) A molecular test based on RT-LAMP for rapid, sensitive and inexpensive colorimetric detection of SARS-CoV-2 in clinical samples. Sci Rep 11:16430. https://doi.org/10.1038/s41598-021-95799-6
Amorim DS (1997) Elementos básicos de sistemática filogenética. Sociedade Brasileira de Entomologia, São Paulo
Andersen JC, Wagner DL (2016) Systematics and biological control. In: Van Driesche RG, Simberloff D, Blossey B, Causton C, Hoddle M, Marks CO, Heinz KM, Wagner DL, Warner KD (eds) Integrating biological control into conservation practice. Wiley, Chichester, pp 105–129
Andersen JC, Oboyski P, Davies N, Charlat S, Ewing C, Meyer C, Krehenwinkel H, Lim JY, Noriyuki S, Ramage T, Gillespie RG, Roderick GK (2019) Categorization of species as native or nonnative using DNA sequence signatures without a complete reference library. Ecol Appl 29(5):e01914. https://doi.org/10.1002/eap.1914
Andow DA (1991) Vegetational diversity and arthropod population response. Annu Rev Entomol 36:561586. https://doi.org/10.1146/annurev.en.36.010191.003021
Bale J (2011) Harmonization of regulations for invertebrate biocontrol agents in Europe: progress, problems and solutions. J Appl Entomol 135:503–513. https://doi.org/10.1111/j.1439-0418.2011.01611.x
Barbosa P (ed) (1998) Conservation biological control. Academic Press, San Diego
Barratt BIP, Evans AA, Ferguson CM, Barker GM, McNeill MR, Phillips CB (1997) Laboratory nontarget host range of the introduced parasitoids Microctonus aethiopoides and M. hyperodae (Hymenoptera: Braconidae) compared with field parasitism in New Zealand. Environ Entomol 26:694–702. https://doi.org/10.1093/ee/26.3.694
Barratt BIP, Blossey B, Hokkanen HM (2006) Post-release evaluation of non-target effects of biological control agents. In: Babendreier D, Bigler F, Kuhlmann U (eds) Environmental impact of invertebrates for biological control of arthropods: Methods and risk assessment. CAB International, Wallingford, pp 166–186
Barratt BIP, Howarth FG, Withers TM, Kean JM, Ridley GS (2010) Progress in risk assessment for classical biological control. Biol Control 52:245–254. https://doi.org/10.1016/j.biocontrol.2009.02.012
Barratt BIP, Moran VC, Bigler F, van Lenteren JC (2018a) The status of biological control and recommendations for improving uptake for the future. Biocontrol 63:155–167. https://doi.org/10.1007/s10526-017-9831-y
Barratt BIP, Cock MJ, Oberprieler RG (2018b) Weevils as targets for biological control, and the importance of taxonomy and phylogeny for efficacy and biosafety. Diversity 10(3):73. https://doi.org/10.3390/d10030073
Basse B, Phillips CB, Hardwick S, Kean JM (2015) Economic benefits of biological control of Sitona obsoletus (clover root weevil) in Southland pasture. N Z Plant Prot 68:218–226. https://doi.org/10.30843/nzpp.2015.68.5809
Begerow D, Nilsson H, Unterseher M, Maier W (2010) Current state and perspectives of fungal DNA barcoding and rapid identification procedures. Appl Microbiol Biotechnol 87:99–108. https://doi.org/10.1007/s00253-010-2585-4
Begg GS, Cook SM, Dye R, Ferrante M, Franck P, Lavigne C, Lövei GL, Mansion-Vaquie A, Pell JK, Petit S, Quesada N (2017) A functional overview of conservation biological control. Crop Prot 97:145–158. https://doi.org/10.1016/j.cropro.2016.11.008
Beltrà A, Addison P, Ávalos JA, Crochard D, Garcia-Marí F, Guerrieri E, Giliomee JH, Malausa T, Navarro-Campos C, Palero F, Soto A (2015) Guiding classical biological control of an invasive mealybug using integrative taxonomy. PLoS One 10(6):e0128685. https://doi.org/10.1371/journal.pone.0128685
Bickford D, Lohman DJ, Sodhi NS, Ng PK, Meier R, Winker K, Ingram KK, Das I (2007) Cryptic species as a window on diversity and conservation. Trends Ecol Evol 22:148–155. https://doi.org/10.1016/j.tree.2006.11.004
Bidartondo MI, Bruns TD, Blackwell M, Edwards I, Taylor AF, Horton T, Zhang N, Koljalg U, May G, Kuyper TW et al (2008) Preserving accuracy in GenBank. Science 319(5870):1616. https://doi.org/10.1126/science.319.5870.1616a
Bin F, Roversi P, van Lenteren J (2012) Erroneous host identification frustrates systematics and delays implementation of biological control. Redia 95:83–88
Blacket MJ, Agarwal A, Zheng L, Cunningham JP, Britton D, Schneider I, Rodoni BC (2020) A LAMP assay for the detection of Bactrocera tryoni Queensland fruit fly (Diptera: Tephritidae). Sci Rep 10:9554. https://doi.org/10.1038/s41598-020-65715-5
Blaimer BB, Lloyd MW, Guillory WX, Brady SG (2016) Sequence capture and phylogenetic utility of genomic ultraconserved elements obtained from pinned insect specimens. PLoS One 11:e0161531. https://doi.org/10.1186/s12862-015-0552-5
Blaser S, Diem H, von Felten A, Gueuning M, Andreou M, Boonham N, Tomlinson J, Müller P, Utzinger J, Frey JE, Bühlmann A (2018) From laboratory to point of entry: development and implementation of a loop-mediated isothermal amplification (LAMP)-based genetic identification system to prevent introduction of quarantine insect species. Pest Manag Sci 74:1504–1512. https://doi.org/10.1002/ps.4866
Boivin G, Kölliker-Ott UM, Bale JS, Bigler F (2006) Assessing the establishment potential of inundative biological control agents. In: Bigler F, Babendreier D, Kuhlmann U (eds) Environmental impact of invertebrates for biological control of arthropods: methods and risk assessment. CAB International, Wallingford, pp 98–113
BOLD (2022) Barcode of Life Database Systems. http://boldsystems.org. Accessed 27 May 2022.
Borghuis A, Pinto JD, Platner GR, Stouthamer R (2004) Partial cytochrome oxidase II sequences distinguish the sibling species Trichogramma minutum Riley and Trichogramma platneri Nagarkatti. Biol Control 30:90–94. https://doi.org/10.1016/j.biocontrol.2003.09.015
Borisenko AV, Sones JE, Hebert PDN (2009) The front-end logistics of DNA barcoding: challenges and prospects. Mol Ecol Resour 9:27–34. https://doi.org/10.1111/j.1755-0998.2009.02629.x
Bortolus A (2008) Error cascades in the biological sciences: the unwanted consequences of using bad taxonomy in ecology. Ambio 37:114–118. https://doi.org/10.1579/0044-7447(2008)37[114:ECITBS]2.0.CO;2
Bridge PD, Roberts PJ, Spooner BM, Panchal G (2003) On the unreliability of published DNA sequences. New Phytol 160:43–48. https://doi.org/10.1046/j.1469-8137.2003.00861.x
Buffington ML, Talamas EJ, Hoelmer KA (2018) Team Trissolcus: integrating taxonomy and biological control to combat the brown marmorated stink bug. Am Entomol 64:224–232. https://doi.org/10.1093/ae/tmy057
Cameron S, Rubinoff D, Will K (2006) Who will actually use DNA barcoding and what will it cost? Syst Biol 55:844–847. https://doi.org/10.1080/10635150600960079
Carson R (1962) Silent Spring. Houghton Mifflin, Boston
Carvalho MR, Bockmann FA, Amorim DS, Vivo M, Toledo-Piza M, Menezes NA, Figueiredo JL, Castro RM, Gill AC, McEachran JD, Compagno LJ (2005) Revisiting the taxonomic impediment. Science 307:353. https://doi.org/10.1126/science.307.5708.353b
Carvalho MR, Bockmann FA, Amorim DS, Brandão CR, Vivo M, Figueiredo JL, Britski HA, Pinna MC, Menezes NA, Marques FP, Papavero N (2007) Taxonomic impediment or impediment to taxonomy? A commentary on systematics and the cybertaxonomic-automation paradigm. Evol Biol 34:140–143. https://doi.org/10.1007/s11692-007-9011-6
CBD (Convention on Biological Diversity) (1992) Convention on biological diversity. https://www.cbd.int/. Accessed 22 January 2020
CBD (Convention on Biological Diversity) (2011) Nagoya Protocol on access and benefit-sharing. Secretariat of the Convention on Biological Diversity, Québec. https://www.cbd.int/abs/. Accessed: 8 Dec 2022
Clarke AR (1993) A new Trissolcus Ashmead species (Hymenoptera: Scelionidae) from Pakistan: species description and its role as a biological control agent. Bull Entomol Res 83:523–527. https://doi.org/10.1017/S0007485300039948
Clausen CP (1942) The relation of taxonomy to biological control. Econ Entomol 35:744–748. https://doi.org/10.1093/jee/35.5.744
Cock MJ, van Lenteren JC, Brodeur J, Barratt BI, Bigler F, Bolckmans K, Cônsoli FL, Haas F, Mason PG, Parra JRP (2010) Do new access and benefit sharing procedures under the convention on biological diversity threaten the future of biological control? Biocontrol 55:199–218. https://doi.org/10.1007/s10526-009-9234-9
Cock MJ, Murphy ST, Kairo MT, Thompson E, Murphy RJ, Francis AW (2016) Trends in the classical biological control of insect pests by insects: an update of the BIOCAT database. Biocontrol 61:349–363. https://doi.org/10.1007/s10526-016-9726-3
Coissac E, Hollingsworth PM, Lavergne S, Taberlet P (2016) From barcodes to genomes: extending the concept of DNA barcoding. Mol Ecol 25:1423–1428. https://doi.org/10.1111/mec.13549
Collier T, Van Steenwyk R (2004) A critical evaluation of augmentative biological control. Biol Control 31:245–256. https://doi.org/10.1016/j.biocontrol.2004.05.001
Collins RA, Cruickshank RH (2013) The seven deadly sins of DNA barcoding. Mol Ecol Resour 13:969–975. https://doi.org/10.1111/1755-0998.12046
Compere H (1969) The role of systematics in biological control: a backward look. Israel J Entomol 4:5–10
Conti E, Avila G, Barratt B, Cingolani F, Colazza S, Guarino S, Hoelmer K, Laumann RA, Maistrello L, Martel G et al (2021) Biological control of invasive stink bugs: review of global state and future prospects. Entomol Exp Appl 169:28–51. https://doi.org/10.1111/eea.12967
Coutinot D, Briano J, Parra JRP, De Sá LAN, Cônsoli FL (2013) Exchange of natural enemies for biological control: is it a rocky road?—The road in the Euro-Mediterranean Region and the South American Common Market. Neotrop Entomol 42:1–14. https://doi.org/10.1007/s13744-012-0103-3
CropLife Brasil (2022) Mercado de biodefensivos no Brasil. https://croplifebrasil.org/. Accessed 04 June 2022
Cruaud A, Groussier G, Genson G, Sauné L, Polaszek A, Rasplus JY (2018) Pushing the limits of whole genome amplification: successful sequencing of RADseq library from a single microhymenopteran (Chalcidoidea, Trichogramma). PeerJ 6:e5640 (https://peerj.com/articles/5640/)
Cruaud A, Nidelet S, Arnal P, Weber A, Fusu L, Gumovsky A, Huber J, Polaszek A, Rasplus J-y (2019) Optimized DNA extraction and library preparation for minute arthropods: application to target enrichment in chalcid wasps used for biocontrol. Mol Ecol Resour 19:702–710. https://doi.org/10.1111/1755-0998.13006
Dang XL, Wen SY, He XF, Pang XF (2005) M-PCR: a powerful method for rapid molecular identification of Trichogramma wasps. Insect Sci 12:77–85. https://doi.org/10.1111/j.1744-7917.2005.00009.x
Danks HV (1988) Systematics in support of entomology. Annu Rev Entomol 33:271–294
Davies AP, Lange CL, O’Neill SL (2006) A rapid single-step multiplex method for discriminating between Trichogramma species in Australia. J Econ Entomol 99:2142–2145. https://doi.org/10.1093/jee/99.6.2142
Dayrat B (2005) Towards integrative taxonomy. Biol J Linn Soc Lond 85:407–415. https://doi.org/10.1111/j.1095-8312.2005.00503.x
de León JH, Jones WA, Morgan DJ (2004) Molecular distinction between populations of Gonatocerus morrilli, egg parasitoids of the glassy-winged sharpshooter from Texas and California: do cryptic species exist? J Insect Sci 4:39. https://doi.org/10.1093/jis/4.1.39
de León JH, Jones WA, Sétamou M, Morgan DJ (2006) Genetic and hybridization evidence confirms that a geographic population of Gonatocerus morrilli (Hymenoptera: Mymaridae) from California is a new species: egg parasitoids of the glassy-winged sharpshooter Homalodisca coagulata (Homoptera: Cicadellidae). Biol Control 38:282–293. https://doi.org/10.1016/j.biocontrol.2006.03.005
DeBach P (1960) The importance of taxonomy to biological control as illustrated by the cryptic history of Aphytis holoxanthus n. sp. (Hymenoptera: Aphelinidae), a parasite of Chrysomphalus aonidum, and Aphytis coheni n. sp., a parasite of Aonidiella aurantii. Ann Entomol Soc Am 53(6):701–705. https://doi.org/10.1093/aesa/53.6.701
DeBach P, Hagen KS (1964) Manipulation of entomophagous species. In: DeBach P (ed) Biological control of insect pests and weeds. Chapman and Hall, London, pp 429–458
DeBach P, Rosen D (1991) Biological control by natural enemies, 2nd edn. Cambridge University Press, Cambridge
Decaux F (1899) Destruction rationnelle des insectes qui attaquent les arbres fruitiers par l’emploi simultané des insecticides, des insectes auxiliaires, et par la protection et l’elevage de leurs ennemis naturelles parasites. J Soc Hort Fr 22:158–184
del Pino M, Rugman-Jones P, Hernández-Suárez E, Polaszek A, Stouthamer R (2013) Rapid molecular identification of five species of Trichogramma occurring in the Canary Islands with notes on their distribution in banana groves. Biocontrol 58:515–524. https://doi.org/10.1007/s10526-013-9519-x
Delucchi V, Rosen D, Schlinger EI (1976) Relationship of systematics to biological control. In: Huffaker CB, Messenger PS (eds) Theory and practice of biological control. Academic Press, New York, pp 81–91
Dermauw W, Van Moerkerke Y, Ebrahimi N, Casteels H, Bonte J, Witters J (2022) A loop-mediated isothermal amplification (LAMP) assay for rapid identification of Ceratitis capitata and related species. Curr Res Insect Sci 2:100029. https://doi.org/10.1016/j.cris.2022.100029
Derocles SA, Plantegenest M, Rasplus JY, Marie A, Evans DM, Lunt DH, Le Ralec A (2016) Are generalist Aphidiinae (Hym. Braconidae) mostly cryptic species complexes? Syst Entomol 41:379–391. https://doi.org/10.1111/syen.12160
DeSalle R, Egan MG, Siddall M (2005) The unholy trinity: taxonomy, species delimitation and DNA barcoding. Philos Trans R Soc Lond B Biol Sci 360(1462):1905–1916. https://doi.org/10.1098/rstb.2005.1722
Dios RDVP, Nihei SS (2020) Taxonomic revision of the genus Trichopoda Berthold, 1827 (Diptera: Tachinidae: Phasiinae), with emphasis on the Neotropical fauna. Zootaxa 4870:1–104. https://doi.org/10.11646/zootaxa.4870.1.1
Dios RDVP, Ziegler J, Zeegers T (2021) The American genus Trichopoda (Diptera: Tachinidae) in Europe–Decades of a misidentified invasive species). Contrib Entomol 71:221–225. https://doi.org/10.21248/contrib.entomol.71.2.221-225
Drew LW (2011) Are we losing the science of taxonomy? As need grows, numbers and training are failing to keep up. Bioscience 61:942–946. https://doi.org/10.1016/j.cris.2022.100029
Eilenberg J, Hajek A, Lomer C (2001) Suggestions for unifying the terminology in biological control. Biocontrol 46:387–400. https://doi.org/10.1023/A:1014193329979
Engel MS, Ceríaco LM, Daniel GM, Dellapé PM, Löbl I, Marinov M, Reis RE, Young MT, Dubois A, Agarwal I, Lehmann AP et al (2021) The taxonomic impediment: a shortage of taxonomists, not the lack of technical approaches. Zool J Linn Soc 193(2):381–387. https://doi.org/10.1093/zoolinnean/zlab072
FAO (Food and Agriculture Organization of the United Nations) (1996) International Standards for Phytosanitary Measures (ISPM), No. 3: Code of Conduct for the Import and Release of Exotic Biological Control Agents. International Plant Protection Convention (IPPC) Secretariat, Rome, 19 pp. https://www.ippc.int/en/publications/600/. Accessed 23 January 2020
Ferguson CM, Roberts GM, Barratt BIP, Evans AA (1994) The distribution of the parasitoid Microctonus aethiopoides Loan (Hymenoptera: Braconidae) in southern South Island Sitona discoideus Gyllenhal (Coleoptera: Curculionidae) populations. In: Popay AJ (ed) Proceedings of the 47th New Zealand Plant Protection Conference. New Zealand Plant Protection Society Inc., Pahia, pp 261–265
Fontes EMG, Pires CSS, Sujii ER (2020) Estratégias de uso e histórico. In: Fontes EMG, Valadares-Inglis MC (eds) Controle biológico de pragas da agricultura. Embrapa, Brasília, pp 21–44
Fontúrbel FE, Murúa MM, Vieli L (2021) Invasion dynamics of the European bumblebee Bombus terrestris in the southern part of South America. Sci Rep 11:15306. https://doi.org/10.1038/s41598-021-94898-8
Forbes AA, Bagley RK, Beer MA, Hippee AC, Widmayer HA (2018) Quantifying the unquantifiable: why Hymenoptera, not Coleoptera, is the most speciose animal order. BMC Ecol 18:1–11. https://doi.org/10.1186/s12898-018-0176-x
Gagné RJ (1995) Contarinia maculipennis (Diptera: Cecidomyiidae), a polyphagous pest newly reported for North America. Bull Entomol Res 85:209–214. https://doi.org/10.1017/S0007485300034283
Gariepy TD, Kuhlmann U, Haye T, Gillott C, Erlandson M (2005) A simple-step multiplex PCR assay for the detection of European Peristenus spp., parasitoids of Lygus spp. Biocontrol Sci Technol 15:481–495. https://doi.org/10.1080/09583150500086771
Gaston KJ, Mound LA (1993) Taxonomy, hypothesis testing and the biodiversity crisis. Proc R Soc Lond B Biol Sci 251(1331):139–142. https://doi.org/10.1098/rspb.1993.0020
Gay H (2012) Before and after Silent Spring: from chemical pesticides to biological control and integrated pest management — Britain, 1945–1980. Ambix 59(2):88–108. https://doi.org/10.1179/174582312X13345259995930
Gebiola M, Monti MM, Johnson RC, Woolley JB, Hunter MS, Giorgini M, Pedata PA (2016a) A revision of the Encarsia pergandiella species complex (Hymenoptera: Aphelinidae) shows cryptic diversity in parasitoids of whitefly pests. Syst Entomol 42:31–59. https://doi.org/10.1111/syen.12187
Gebiola M, White JA, Cass BN, Kozuch A, Harris LR, Kelly SE, Karimi J, Giorgini M, Perlman SJ, Hunter MS (2016b) Cryptic diversity, reproductive isolation and cytoplasmic incompatibility in a classic biological control success story. Biol J Linn Soc Lond 117:217–230. https://doi.org/10.1111/bij.12648
Gerard PJ, Wilson DJ, Eden TM (2011) Field release, establishment and initial dispersal of Irish Microctonus aethiopoides in Sitona lepidus populations in northern New Zealand pastures. Biocontrol 56:861–870. https://doi.org/10.1007/s10526-011-9367-5
Gillespie MA, Gurr GM, Wratten SD (2016) Beyond nectar provision: the other resource requirements of parasitoid biological control agents. Entomol Exp Appl 159(2):207–221. https://doi.org/10.1111/eea.12424
Gliessman SR (2005) Agroecologia – processos ecológicos em agricultura sustentável. Editora da Universidade Federal do Rio Grande do Sul, Porto Alegre
Godfray HCJ (2002) Challenges for taxonomy. Nature 417:17–19. https://doi.org/10.1038/417017a
Gokhman VE (2018) Integrative taxonomy and its implications for species-level systematics of parasitoid Hymenoptera. Entomol Rev 98:834–864. https://doi.org/10.1134/S0013873818070059
Goldson SL, Proffitt JR, McNeill MR (1990) Seasonal biology and ecology in New Zealand of Microctonus aethiopoides (Hymenoptera: Braconidae), a parasitoid of Sitona spp. (Coleoptera: Curculionidae), with special emphasis on atypical behaviour. J Appl Ecol 27:703–722. https://doi.org/10.2307/2404313
Goldson SL, McNeill MR, Proffitt JR (2003) Negative effects of strain hybridisation on the biocontrol agent Microctonus aethiopoides. N Z Plant Prot 56:138–142. https://doi.org/10.30843/nzpp.2003.56.6055
Goldson SL, McNeill MR, Proffitt JR, Barratt BP (2005) Host specificity testing and suitability of a European biotype of the braconid parasitoid Microctonus aethiopoides as a biological control agent against Sitona lepidus (Coleoptera: Curculionidae) in New Zealand. Biocontrol Sci Technol 15:791–813. https://doi.org/10.1080/09583150500136444
Gonçalves DDS, Cassimiro APA, de Oliveira CD, Rodrigues NB, Moreira LA (2014) Wolbachia detection in insects through LAMP: loop mediated isothermal amplification. Parasit Vectors 7:1–5. https://doi.org/10.1186/1756-3305-7-228
Gonzales-Cabrera J, Arredondo-Bernal HC, Stouthamer R (2014) Multiplex PCR assay to identify Trichogramma parasitoids (Hymenoptera: Trichogrammatidae) reared from Mexican insectaries. Agrociencia 48:703–711
González-Hernández A, López-Arrollo JI (2007) Importancia de la sistemática en control biológico. In: Arredondo Bernal HC (ed) Rodríguez del Bosque LA. Teoría y aplicación del control biológico. Sociedad Mexicana de Control Biológico, Mexico City, pp 36–47
Gordh G (1977) Biosystematics of natural enemies. In: Ridgway RL, Vinson SB (eds) Biological control by augmentation of natural enemies. Plenum, New York, pp 125–148
Gordh G, Beardsley JW (1999) Taxonomy and biological control. In: Bellows TS, Fisher TW (eds) Handbook of biological control. Academic Press, San Diego, pp 45–55
Goulet H, Mason PG (2006) Review of the Nearctic species of Leiophron and Peristenus (Hymenoptera: Braconidae: Euphorinae) parasitizing Lygus (Hemiptera: Miridae: Mirini). Zootaxa 1323:1–118. https://doi.org/10.11646/zootaxa.1323.1.1
Grant DM, Brodnicke OB, Evankow AM et al (2021) The future of DNA barcoding: reflections from early career researchers. Diversity 13:313. https://doi.org/10.3390/d13070313
Grissell EE (1999) Hymenopteran biodiversity: some alien notions. Am Entomol 45:235–244. https://doi.org/10.1093/ae/45.4.235
Gurr GM, Wratten SD, Altieri MA (2004) Ecological engineering: a new direction for agricultural pest management. Aust Farm Bus Manag J 1:28–35
Hajek AE, Hurley BP, Kenis M, Garnas JR, Bush SJ, Wingfield MJ, van Lenteren JC, Cock MJ (2016) Exotic biological control agents: a solution or contribution to arthropod invasions? Biol Invasions 18:953–969. https://doi.org/10.1007/s10530-016-1075-8
Hanson PE (1993) La importancia de la taxonomía en el control biológico. Manejo Integrado De Plagas 29:48–50
Hardy DE, Delfinado MD (1980) Insects of Hawaii, vol. 13. Diptera: Cyclorrhapha III. University of Hawaii Press, Honolulu
Hawkins BA, Lawton JH (1987) Species richness for parasitoid of British polyphagous insects. Nature 326:788–790
Hebert PD, Cywinska A, Ball SL, DeWaard JR (2003a) Biological identifications through DNA barcodes. Proc R Soc Lond B 270(1512):313–321. https://doi.org/10.1098/rspb.2002.2218
Hebert PD, Ratnasingham S, De Waard JR (2003b) Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond B 270(suppl):S96–S99. https://doi.org/10.1098/rsbl.2003.0025
Heimpel GE, Cock MJ (2018) Shifting paradigms in the history of classical biological control. Biocontrol 63:27–37. https://doi.org/10.1007/s10526-017-9841-9
Hennig W (1999) Phylogenetic systematics. University of Illinois Press, Urbana
Heraty JM (1998) Systematics: science or service? In: Innovations in biological control research. Proceedings, California Conference on Biological Control I. University of California, Berkeley, pp 187–190
Heraty JM (2004) Molecular systematics, Chalcidoidea and biological control. In: Ehler LE, Siorza R, Mateille T (eds) Genetics, evolution and biological control. CAB International, London, pp 39–71
Heraty J (2009) Parasitoid biodiversity and insect pest management. In: Foottit RG, Adler PH (eds) Insect biodiversity: science and society. Wiley-Blackwell, Oxford, pp 445–462
Heraty JM, Polaszek A (2000) Morphometric analysis and descriptions of selected species in the Encarsia strenua group (Hymenoptera: Aphelinidae). J Hymenopt Res 9:142–169
Heraty JM, Woolley JB, Hopper KM, Hawks DL, Kim J-W, Buffington M (2007) Phylogenetic relationships of cryptic species in the Aphelinus varipes complex (Hymenoptera: Aphelinidae). Mol Phylogenet Evol 45:480–493. https://doi.org/10.1016/j.ympev.2007.06.021
Hickerson MJ, Meyer CP, Moritz C (2006) DNA barcoding will often fail to discover new animal species over broad parameter space. Syst Biol 55:729–739. https://doi.org/10.1080/10635150600969898
Hita-Garcia F, Lieberman Z, Audisio TL, Liu C, Economo EP (2019) Revision of the highly specialized ant genus Discothyrea (Hymenoptera: Formicidae) in the Afrotropics with X-ray microtomography and 3D cybertaxonomy. Insect Syst Divers 3(6):5. https://doi.org/10.1093/isd/ixz015
Hoddle MS (2004) Analysis of fauna in the receiving area for the purpose of identifying native species that exotic natural enemies may potentially attack. In: Van Driesche RG, Reardon R (eds) Assessing host ranges for parasitoids and predators used for classical biological control: a guide to best practice. Forest Health Technology Enterprise Team USDA Forest Service, Morgantown, pp 24–39
Hopper KR, Britch SC, Wajnberg E (2006) Risks of interbreeding between species used in biological control and native species, and methods for evaluating their occurrence and impact. In: Bigler F, Babendreier D, Kuhlmann U (eds) Environmental impact of invertebrates for biological control of arthropods: methods and risk assessment. CABI Publishing, Delémont, pp 78–97
Howarth FG (1983) Classical biological control: panacea or Pandora’s box. Proc Hawaii Entomol Soc 24:239–244
Howarth FG (1991) Environmental impacts of classical biological control. Annu Rev Entomol 36:489–509
Howarth FG (2000) Non-target effects of biological control agents. In: Gurr G, Wratten S (eds) Biological control: measures of success. Springer, Dordrecht, pp 369–403
Hsieh CH, Wang HY, Chen YF, Ko CC (2012) Loop-mediated isothermal amplification for rapid identification of biotypes B and Q of the globally invasive pest Bemisia tabaci, and studying population dynamics. Pest Manag Sci 68(8):1206–1213. https://doi.org/10.1002/ps.3298
Huang JP (2020) Is population subdivision different from speciation? From phylogeography to species delimitation. Ecol Evol 10:6890–6896. https://doi.org/10.1002/ece3.6524
Huber JT (2015) World reclassification of the Gonatocerus group of genera (Hymenoptera: Mymaridae). Zootaxa 3967(1):1–84. https://doi.org/10.11646/zootaxa.3967.1.1
Huber JT, Liu M, Fernández-Triana J (2002) Taxonomy and biological control. In: Mason PG, Huber JT (eds) Biological control programmes in Canada, 1981–2000. CABI Publishing, Ottawa, pp 14–22
Huber JT, Liu M, Fernández-Triana J (2021) Taxonomy and biological control. In: Mason PG (ed) Biological control: global impacts, challenges and future directions of pest management. CSIRO Publishing, Boca Raton, pp 236–260
Hubert N, Hanner R (2015) DNA barcoding, species delineation and taxonomy: a historical perspective. DNA Barcodes 3:44–58. https://doi.org/10.1515/dna-2015-0006
Hunt EJ, Kuhlmann U, Sheppard A, Qin TK, Barratt BIP, Harrison L, Mason PG, Parker D, Flanders RV, Goolsby J (2008) Review of invertebrate biological control agent regulation in Australia, New Zealand, Canada and the USA: recommendations for a harmonized European system. J Appl Entomol 132:89–123. https://doi.org/10.1111/j.1439-0418.2007.01232.x
Hunter WD (1901) The Aphididae of North America. Iowa Agric Coll Exp Stn Bull 60:63–138
Ilharco FA, van Harten A (1987) Systematics. In: Minks AK, Harrewijn P (eds) Aphids: their biology, natural enemies and control, Vol. A. Elsevier (World Crop Pests, 2A), Amsterdam, pp 51–77
IBAMA (Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis) (2016) Instrução Normativa IN no 5, de agosto de 2016.
ISSG (Invasive Species Specialist Group) (2018) The application of biological control for the management of established invasive alien species causing environmental impacts. Sheppard AW, Paynter Q, Mason P, Murphy S, Stoett P, Cowan P, Brodeur J, Warner K, Villegas C, Shaw R, Hinz H, Hill M, Genovesi P (eds) Sharm El-Sheikh: Convention on Biological Diversity (CBD) information document for the convention on biological diversity, 88 pp. Available: https://www.cbd.int/doc/c/0c6f/7a35/eb8815eff54c3bc4a02139fd/cop-14-inf-09-en.pdf. Accessed 23 January 2020
Ivezić A, Rugman-Jones PF, Stouthamer R, Ignjatović-Ćupina A (2018) Molecular identification of Trichogramma egg parasitoids of Ostrinia nubilalis in northeastern Serbia. Arch Biol Sci 70:425–432. https://doi.org/10.2298/ABS171103002I
Ivezić A, Rugman-Jones PF, Thibaut M, Ris N, Ignjatović-Ćupina A (2021) Molecular identification of Trichogramma species parasitizing Ostrinia nubilalis in corn and pepper in south-east border of Europe. Int J Pest Manag 67:346–357. https://doi.org/10.1080/09670874.2020.1779383
Jenkins C, Chapman TA, Micallef JL, Reynolds OL (2012) Molecular techniques for the detection and differentiation of host and parasitoid species and the implications for fruit fly management. Insects 3:763–788. https://doi.org/10.3390/insects3030763
Joly S, Davies TJ, Archambault A, Bruneau A, Derry A, Kembel SW, Peres-Neto P, Vamosi J, Wheeler TA (2014) Ecology in the age of DNA barcoding: the resource, the promise and the challenges ahead. Mol Ecol Resour 14:221–232. https://doi.org/10.1111/1755-0998.12173
Kairo MT, Cock MJ, Quinlan MM (2003) An assessment of the use of the Code of Conduct for the Import and Release of Exotic Biological Control Agents (ISPM No. 3) since its endorsement as an international standard. Biocontrol News and Information 24:15N-27N. https://doi.org/10.1079/cabireviews20083023401
Kim KC, McPheron BA (1993) Biology of variation: epilogue. In: Kim KC, McPheron BA (eds) Evolution of insect pests: patterns of variation. John Wiley and Sons, New York, pp 453–468
Kitano D, Takakura KI (2020) Simple and on-site DNA purification for LAMP reaction applicable to non-adult tephritid fruit fly (Diptera: Tephiritidae). J Appl Entomol 144:824–829. https://doi.org/10.1111/jen.12820
Knutson L (1981) Symbiosis of biosystematics and biocontrol. In: Papavizas GC (ed), Biological control in crop production. Beltsville Symposium 5. Allanheld, Osmun, Totowa, pp 61–78
Kuske S, Widmer F, Edwards PJ, Turlings TC, Babendreier D, Bigler F (2003) Dispersal and persistence of mass released Trichogramma brassicae (Hymenoptera: Trichogrammatidae) in non-target habitats. Biol Control 27:181–193. https://doi.org/10.1016/S1049-9644(02)00191-3
Lipscomb D, Platnick N, Wheeler Q (2003) The intellectual content of taxonomy: a comment on DNA taxonomy. Trends Ecol Evol 18(2):65–66. https://doi.org/10.1016/S0169-5347(02)00060-5
Loomans AJ (2021) Every generalist biological control agent requires a special risk assessment. Biocontrol 66:23–35. https://doi.org/10.1007/s10526-020-10022-1
Louda SM, Pemberton RW, Johnson MT, Follett P (2003) Nontarget effects—the Achilles’ heel of biological control? Retrospective analyses to reduce risk associated with biocontrol introductions. Annu Rev Entomol 48:365–396. https://doi.org/10.1146/annurev.ento.48.060402.102800
Lowenstein JH, Amato G, Kolokotronis SO (2009) The real maccoyii: identifying tuna sushi with DNA barcodes – contrasting characteristic attributes and genetic distances. PLoS ONE 4:e7866. https://doi.org/10.1371/journal.pone.0007866
Lynch L, Hokkanen HMT, Babendreier D, Bigler F, Burgio G, Gao ZH, Kuske S, Loomans A, Menzler-Hokkanen I, Thomas MB, Tommasini G (2001) Insect biocontrol and non-target effects: a European perspective. In: Wajnberg E, Scott JC, Quimby PC (eds) Evaluating indirect ecological effects of biological control. CABI Publishing, Wallingford, pp 99–125
Maddox JV, McManus ML, Jeffords MR, Webb RE (1992) Exotic insect pathogens as classical biological control agents with an emphasis on regulatory considerations. In: Kauffman WC, Nechols JE (eds) Selection criteria and ecological consequences of importing natural enemies. Entomological Society of America, Lanham, pp 27–39
Manzari S, Polaszek A, Belshaw R, Quicke DLJ (2002) Morphometric and molecular analysis of the Encarsia inaron species-group (Hymenoptera: Aphelinidae), parasitoids of whiteflies (Hemiptera: Aleyrodidae). Bull Entomol Res 92:165–175. https://doi.org/10.1079/BER2001144
MAPA/ANVISA/IBAMA (Ministério da Agricultura, Pecuária e Abastecimento / Agência Nacional de Vigilância Sanitária / Instituto Brasileiro do Meio Ambiente e dos Recursos Naturais Renováveis) (2019) Instrução Normativa Conjunta de No. 1, de 8 de novembro de 2019.
Marinho CF, Costa VA, Zucchi RA (2018) Annotated checklist and illustrated key to braconid parasitoids (Hymenoptera, Braconidae) of economically important fruit flies (Diptera, Tephritidae) in Brazil. Zootaxa 4527:21–26. https://doi.org/10.11646/zootaxa.4527.1.2
Mason PG, De Clerck-Floate RA, Gallant B, Gillespie DR, Floate K, Bourchier R, Douglas H, Vincent C, Boivin G (2017) Guide for the first-time importation and release of arthropod biological control agents in Canada. Canadian Food Inspection Agency, Ottawa. http:// www.publications.gc.ca/site/eng/9.843006/ publication.html. Accessed 22 January 2020
Mayr E (1958) Concepts of biology. National Academy of Sciences, National Research Council, Publication 560, Washington D.C. 118p.
Mayr E (1968) The role of systematics in biology. Science 159:595–599
Meier R (2017) Citation of taxonomic publications: the why, when, what and what not. Syst Entomol 42:301–304. https://doi.org/10.1111/syen.12215
Meier R, Shiyang K, Vaidya G, Ng PKL (2006) DNA barcoding and taxonomy in Diptera: a tale of high intraspecific variability and low identification success. Syst Biol 55:715–728. https://doi.org/10.1080/10635150600969864
Meier R, Blaimer BB, Buenaventura E, Hartop E, von Rintelen T, Srivathsan A, Yeo D (2022) A re-analysis of the data in Sharkey et al.’s (2021) minimalist revision reveals that BINs do not deserve names, but BOLD Systems needs a stronger commitment to open science. Cladistics 38:264–275. https://doi.org/10.1111/cla.12489
Meiklejohn KA, Damaso N, Robertson JM (2019) Assessment of BOLD and GenBank—their accuracy and reliability for the identification of biological materials. PLoS One 14:e0217084. https://doi.org/10.1371/journal.pone.0217084
Meyer CP, Paulay G (2005) DNA barcoding: error rates based on comprehensive sampling. PLoS Biol 3:2229–2238. https://doi.org/10.1371/journal.pbio.0030422
Miller GL, Foottit RG (2009) The taxonomy of crop pests: the aphids. In: Foottit RG, Adler PH (eds) Insect biodiversity: science and society, 1st edn. Wiley-Blackwell, Oxford, pp 463–473
Miller SE (2007) DNA barcoding and the renaissance of taxonomy. Proc Natl Acad Sci USA 104:4775–4776. https://doi.org/10.1073/pnas.0700466104
Minelli A (2020) Taxonomy needs pluralism, but a controlled and manageable one. Megataxa 001(1):009–018. https://doi.org/10.11646/megataxa.1.1.3
Moraes GJ (1987) Importance of taxonomy in biological control. Int J Trop Insect Sci 8:841–844. https://doi.org/10.1017/S1742758400023031
Murdoch WW, Briggs CJ, Nisbet RM (2003) Consumer-resource dynamics. Monographs in Population Biology 36. Princeton University Press, Princeton
Nam H, Kwon M, Ramasamy S, Kim J (2022) Identification of two diamondback moth parasitoids, Diadegma fenestrale and Diadegma semiclausum, using LAMP for application in biological control. Horticulturae 8:366. https://doi.org/10.3390/horticulturae8050366
Neuenschwander P (2001) Biological control of the cassava mealybug in Africa: a review. Biol Control 21:214–229. https://doi.org/10.1006/bcon.2001.0937
Nilsson RH, Ryberg M, Kristiansson E, Abarenkov K, Larsson KH, Koljalg U (2006) Taxonomic reliability of DNA sequences in public sequence databases: a fungal perspective. PLoS One 1:e59. https://doi.org/10.1371/journal.pone.0000059
Notomi T, Okayama H, Masubuchi H, Yonekawa T, Watanabe K, Amino N, Hase T (2000) Loop-mediated isothermal amplification of DNA. Nucleic Acids Res 28:e63. https://doi.org/10.1093/nar/28.12.e63
Notomi T, Mori Y, Tomita N, Kanda H (2015) Loop-mediated isothermal amplification (LAMP): principle, features, and future prospects. J Microbiol 53:1–5. https://doi.org/10.1007/s12275-015-4656-9
Noyes JS (1994) The reliability of published host-parasitoid records: a taxonomist’s view. Norweg J Agric Sci 16:59–69
Noyes JS (2019) Universal Chalcidoidea Database. World Wide Web electronic publication. http://www.nhm.ac.uk/chalcidoids. Accessed 12 July 2020
Packer L, Gibbs J, Sheffield C, Hanner R (2009) DNA barcoding and the mediocrity of morphology. Mol Ecol Resour 9:42–50. https://doi.org/10.1111/j.1755-0998.2009.02631.x
Packer L, Monckton SK, Onuferko TM, Ferrari RR (2018) Validating taxonomic identifications in entomological research. Insect Conserv Divers 11:1–12. https://doi.org/10.1111/icad.12284
Parra JRP (2014) Biological control in Brazil: an overview. Sci Agric 71:420–429. https://doi.org/10.1590/0103-9016-2014-0167
Parra JRP, Coelho A Jr (2019) Applied biological control in Brazil: from laboratory assays to field application. J Insect Sci 19:1–6. https://doi.org/10.1093/jisesa/iey112
Parra JRP, Botelho PSM, Corrêa-Ferreira B, Bento JMS (2002) Controle biológico no Brasil: parasitóides e predadores. Manole, São Paulo
Parra JRP, Pinto AS, Nava DE, Oliveira RC, Diniz AJF (2021) Conceitos e evolução do controle biológico. In: Parra JRP, Pinto AS, Nava DE, Oliveira RC, Diniz AJF (eds) Controle biológico no Brasil com parasitoides e predadores na agricultura brasileira. Fundação de Estudos Agrários Luiz de Queiroz, Piracicaba, pp 17–39
Peixoto L, Allen GR, Ridenbaugh RD, Quarrell SR, Withers TM, Sharanowski BJ (2018) When taxonomy and biological control researchers unite: Species delimitation of Eadya parasitoids (Braconidae) and consequences for classical biological control of invasive paropsine pests of Eucalyptus. PLoS One 13(8):e0201276. https://doi.org/10.1371/journal.pone.0201276
Phillips CB, Vink CJ, Blanchet A, Hoelmer KA (2008) Hosts are more important than destinations: what genetic variation in Microctonus aethiopoides (Hymenoptera: Braconidae) means for foreign exploration for natural enemies. Mol Phylogenet Evol 49:467–476. https://doi.org/10.1016/j.ympev.2008.08.005
Piffaretti J, Vanlerberghe-Masutti F, Tayeh A, Clamens AL, D’Acier AC, Jousselin E (2012) Molecular phylogeny reveals the existence of two sibling species in the aphid pest Brachycaudus helichrysi (Hemiptera: Aphididae). Zool Scr 41(3):266–280. https://doi.org/10.1111/j.1463-6409.2012.00531.x
Pimentel D, McNair S, Janecka J, Wightman J, Simmonds C, O’Connell C, Wong E, Russel L, Zern J, Aquino T, Tsomondo T (2001) Economic and environmental threats of alien plant, animal, and microbe invasions. Agric Ecosyst Environ 84:1–20. https://doi.org/10.1016/S0167-8809(00)00178-X
Pinto JD (1999) Systematics of the North American species of Trichogramma Westwood (Hymenoptera: Trichogrammatidae). Mem Entomol Soc Wash 22:1–287
Pinto JD, Koopmanschap AB, Platner GR, Stouthamer R (2002) The North American Trichogramma (Hymenoptera: Trichogrammatidae) parasitizing certain Tortricidae (Lepidoptera) on apple and pear, with ITS2 DNA characterizations and description of a new species. Biol Control 23:134–142. https://doi.org/10.1006/bcon.2001.0995
Pinto JD, Platner GR, Stouthamer R (2003) The systematics of the Trichogramma minutum species complex (Hymenoptera: Trichogrammatidae), a group of important North American biological control agents: the evidence from reproductive compatibility and allozymes. Biol Control 27:167–180. https://doi.org/10.1016/S1049-9644(02)00190-1
Porter TM, Hajibabaei M (2018) Over 2.5 million COI sequences in GenBank and growing. PLoS One 13(9):e0200177. https://doi.org/10.1371/journal.pone.0200177
Qin TK, Gullan PJ, Beattie GAC, Trueman JW, Cranston PS, Fletcher MJ, Sands DP (1994) The current distribution and geographical origin of the scale insect pest Ceroplastes sinensis (Hemiptera: Coccidae). Bull Entomol Res 84:541–549. https://doi.org/10.1017/S000748530003279X
Querino RB, Zucchi RA (2005) An illustrated key to the species of Trichogramma (Hymenoptera: Trichogrammatidae) of Brazil. Zootaxa 1073:37–60. https://doi.org/10.11646/zootaxa.1073.1.3
Querino RB, Zucchi RA (2011) Guia de identificação de Trichogramma para o Brasil. Embrapa Informação Tecnológica, Brasília
Querino RB, Zucchi RA (2019) Annotated checklist and illustrated key to the species of Trichogramma Westwood (Hymenoptera: Trichogrammatidae) from South America. Zootaxa 4656:201–231. https://doi.org/10.11646/zootaxa.4656.2.1
Querino RB, Moraes RCB, Zucchi RA (2002) Relative warp analysis to study morphological variations in the genital capsule of Trichogramma pretiosum Riley (Hymenoptera: Trichogrammatidae). Neotrop Entomol 31:217–224. https://doi.org/10.1590/S1519-566X2002000200007
Querino RB, Zucchi RA, Pinto JD (2010) Systematics of the Trichogrammatidae (Hymenoptera: Chalcidoidea) with a focus on the genera attacking Lepidoptera. In: Consoli FL, Parra JR, Zucchi RA (eds) Egg parasitoids in agroecosystems with emphasis on Trichogramma. Progress in biological control vol. 9. Springer Science and Business Media, Berlin, pp 191–218
Rako L, Agarwal A, Semeraro L, Broadley A, Rodoni BC, Blacket MJ (2021) A LAMP (loop-mediated isothermal amplification) test for rapid identification of Khapra beetle (Trogoderma granarium). Pest Manag Sci 77:5509–5521. https://doi.org/10.1002/ps.6591
Ratnasingham S, Hebert PDN (2007) BOLD: The barcode of life data system (http://www.barcodinglife.org). Mol Ecol Notes 7:355–364. https://doi.org/10.1111/j.1471-8286.2007.01678.x
Raxworthy CJ, Smith BT (2021) Mining museums for historical DNA: advances and challenges in museomics. Trends Ecol Evol 36:1049–1060. https://doi.org/10.1016/j.tree.2021.07.009
Remaudière G, Remaudière M (1997) Catalogue des Aphididae du monde (Homoptera Aphidoidea). INRA Editions, Paris
Ridenbaugh RD, Barbeau E, Sharanowski BJ (2018) Description of four new species of Eadya (Hymenoptera, Braconidae), parasitoids of the Eucalyptus Tortoise Beetle (Paropsis charybdis) and other Eucalyptus defoliating leaf beetles. J Hymenopt Res 64:141–175. https://doi.org/10.3897/jhr.64.24282
Ridgway R, Vinson SB (1977) Biological control by augmentation of natural enemies: insect and mite control with parasites and predators. Plenum Press, New York
Rosen D (1986) The role of taxonomy in effective biological control programs. Agric Ecosyst Environ 15:121–129. https://doi.org/10.1016/0167-8809(86)90085-X
Rosen D, DeBach P (1973) Systematics, morphology and biological control. Entomophaga 18:215–222. https://doi.org/10.1007/BF02371468
Rubinoff D, Cameron S, Will K (2006) A genomic perspective on the shortcomings of mitochondrial DNA for barcoding identification. J Hered 97:581–594. https://doi.org/10.1093/jhered/esl036
Rugman-Jones PF, Forster LD, Guerrieri E, Luck RF, Morse JG, Monti MM, Stouthamer R (2011) Taxon-specific multiplex-PCR for quick, easy, and accurate identification of encyrtid and aphelinid parasitoid species attacking soft scale insects in California citrus groves. Biocontrol 56:265–275. https://doi.org/10.1007/s10526-010-9328-4
Sabahi S, Fekrat L, Zakiaghl M, Moravej GH (2018) Loop-mediated isothermal amplification combined with PCR for rapid identification of the Ethiopian fruit fly (Diptera: Tephritidae). Neotrop Entomol 47:96–105. https://doi.org/10.1007/s13744-017-0522-2
Sabrosky CW (1955) The interrelations of biological control and taxonomy. J Econ Entomol 48:710–714
Santos B, Miller M, Miklasevskaja M, McKeown J, Redmond N, Coddington J, Bird J, Miller S, Smith A, Brady S, Buffington M (2022) Enhancing DNA barcode reference libraries by harvesting terrestrial arthropods at the National Museum of Natural History. ARPHA Preprints 3:e84305. https://doi.org/10.3897/arphapreprints.e84305
Schauff ME, LaSalle J (1998) The relevance of systematics to biological control: protecting the investment in research. In: White GG, Drew RAI, Zalucki MP (eds) Pest management: future challenges, vol 1. Proceedings of the 6th Australian Applied Entomological Conference, Brisbane, pp 425–436
Schlick-Steiner BC, Steiner FM, Seifert B, Stauffer C, Christian E, Crozier RH (2010) Integrative taxonomy: a multisource approach to exploring biodiversity. Annu Rev Entomol 55:421–438. https://doi.org/10.1146/annurev-ento-112408-085432
Schlinger El, Doutt RL (1964) Systematics in relation to biological control. In: DeBach P (ed) Biological control of insect pests and weeds. Chapman and Hall, London, pp 247–280
Schoch CL, Ciufo S, Domrachev M, Hotton CL, Kannan S, Khovanskaya R, Leipe D, Mcveigh R, O’Neill K, Robbertse B et al (2020) NCBI Taxonomy: a comprehensive update on curation resources and tools. Database 2020:baaa062. https://doi.org/10.1093/database/baaa062
Seehausen ML, Ris N, Driss L, Racca A, Girod P, Warot S, Borowiec N, Toševski I, Kenis M (2020) Evidence for a cryptic parasitoid species reveals its suitability as a biological control agent. Sci Rep 10:1–12. https://doi.org/10.1038/s41598-020-76180-5
Sharkey MJ, Fernández F (2006) Biología y diversidade de Hymenoptera. In: Fernández F, Sharkey MJ (eds) Introducción a los Hymenoptera de la región neotropical. Sociedad Colombiana de Entomología y Universidad Nacional de Colombia, Bogotá, pp 93–113
Sharkey MJ, Janzen DH, Hallwachs W, Chapman EG, Smith MA, Dapkey T, Brown A, Ratnasingham S, Naik S, Manjunath R et al (2021) Minimalist revision and description of 403 new species in 11 subfamilies of Costa Rican braconid parasitoid wasps, including host records for 219 species. ZooKeys 1013:1–665. https://doi.org/10.3897/zookeys.1013.55600
Shimbori EM, Costa VA, Zucchi RA (2020) Annotated checklist and illustrated key to parasitoids (Hymenoptera: Diapriidae, Eulophidae and Pteromalidae) of fruit flies (Diptera, Tephritidae) in Brazil. Zootaxa 4858:53–70. https://doi.org/10.11646/zootaxa.4858.1.3
Shimbori EM, Sousa IMM, Domingues VS, Garcia SSP, Nascimento PAG, Penteado-Dias AM (2022) Import and release of invertebrate biological control agents: regulation in Brazil. In: Hokkanen HMT, Menzler-Hokkanen I (eds): Book of Abstracts. 26th International Congress of Entomology, Helsinki
Simberloff D (2009) The role of propagule pressure in biological invasions. Annu Rev Ecol Evol Syst 40:81–102. https://doi.org/10.1146/annurev.ecolsys.110308.120304
Sinkins SP, Braig HR, O’Neill SL (1995) Wolbachia superinfections and the expression of cytoplasmic incompatibility. Proc R Soc Lond B Biol Sci 261:325–330. https://doi.org/10.1098/rspb.1995.0154
Smith-Ramírez C, Vieli L, Barahona-Segovia RM, Montalva J, Cianferoni F, Ruz L, Fontúrbel FE, Valdivia CE, Medel R, Pauchard A et al (2018) Las razones de por qué Chile debe detener la importación del abejorro comercial Bombus terrestris (Linnaeus) y comenzar a controlarlo. Gayana 82(2):118–127
Stigenberg J, Boring CA, Ronquist F (2015) Phylogeny of the parasitic wasp subfamily Euphorinae (Braconidae) and evolution of its host preferences. Syst Entomol 40:570–591. https://doi.org/10.1111/syen.12122
Stork NE (2018) How many species of insects and other terrestrial arthropods are there on Earth? Annu Rev Entomol 63:31–45. https://doi.org/10.1146/annurev-ento-020117-043348
Stouthamer R, Hu J, van Kan FJPM, Platner GR, Pinto JD (1999) The utility of internally transcribed spacer 2 DNA sequences of the nuclear ribosomal gene for distinguishing sibling species of Trichogramma. Biocontrol 43:421–440
Strasser BJ (2008) GenBank – natural history in the 21st century? Science 322(5901):537–538. https://doi.org/10.1023/A:1009937108715
Talamas EJ, Herlihy MV, Dieckhoff C, Hoelmer KA, Buffington ML, Bon M-C, Weber DC (2015) Trissolcus japonicus (Ashmead) (Hymenoptera, Scelionidae) emerges in North America. J Hymenopt Res 43:119–128. https://doi.org/10.3897/JHR.43.8560
Tautz D, Arctander P, Minelli A, Thomas RH, Vogler AP (2003) A plea for DNA taxonomy. Trends Ecol Evol 18:70–74. https://doi.org/10.1016/S0169-5347(02)00041-1
Taylor HR, Harris WE (2012) An emergent science on the brink of irrelevance: a review of the past 8 years of DNA barcoding. Mol Ecol Resour 12:377–388. https://doi.org/10.1111/j.1755-0998.2012.03119.x
Thiele K, Yeates D (2002) Tension arises from duality at the heart of taxonomy. Nature 419(6905):337. https://doi.org/10.1038/419337a
Thomson SA, Pyle RL, Ahyong ST, Alonso-Zarazaga M, Ammirati J, Araya JF, Ascher JS, Audisio TL, Azevedo-Santos VM, Bailly N, Baker WJ (2018) Taxonomy based on science is necessary for global conservation. PLoS Biol 16(3):e2005075. https://doi.org/10.1371/journal.pbio.2005075
Tracy EF (2014) The promise of biological control for sustainable agriculture: a stakeholder-based analysis. J Sci Policy Gov 5(1):1–13.
Triapitsyn SV (2006) A key to the Mymaridae (Hymenoptera) egg parasitoids of proconiine sharpshooters (Hemiptera: Cicadellidae) in the Nearctic region, with description of two new species of Gonatocerus. Zootaxa 1203(1):1–38. https://doi.org/10.11646/zootaxa.1203.1.1
Triapitsyn SV, Huber JT, Logarzo GA, Berezovskiy VV, Aquino DA (2010) Review of Gonatocerus (Hymenoptera: Mymaridae) in the Neotropical region, with description of eleven new species. Zootaxa 2456(1):1–243. https://doi.org/10.11646/zootaxa.2456.1.1
Uechi N, Tokuda M, Yukawa J, Kawamura F, Teramoto KK, Harris KM (2003) Confirmation by DNA analysis that Contarinia maculipennis (Diptera: Cecidomyiidae) is a polyphagous pest of orchids and other unrelated cultivated plants. Bull Entomol Res 93:545–551. https://doi.org/10.1079/BER2003266
Van Driesche R, Bellows TS Jr (1996) Biological control. Springer Science and Business Media, Berlin
Van Driesche R, Hoddle MS (2016) Non-target effects of insect biocontrol agents and trends in host specificity since 1985. CAB Rev 11(44):1–66. https://doi.org/10.1079/PAVSNNR201611044
Van Driesche R, Simberloff D, Blossey B, Causton C, Hoddle M, Marks CO, Heinz KM, Wagner DL, Warner KD (eds) (2016) Integrating biological control into conservation practice. John Wiley and Sons, Chichester
Van Houdt JKJ, Breman FC, Virgilio M, De Meyer M (2010) Recovering full DNA barcodes from natural history collections of Tephritid fruitflies (Tephritidae, Diptera) using mini barcodes. Mol Ecol Resour 10:459–465. https://doi.org/10.1111/j.1755-0998.2009.02800.x
Van Lenteren JC (2012) The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. Biocontrol 57:1–20. https://doi.org/10.1007/s10526-011-9395-1
Velthuis HH, Van Doorn A (2006) A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 37:421–451. https://doi.org/10.1051/apido:2006019
Venzon M, Neves WS, Júnior TJP, Pallini A (2021) Controle alternativo de pragas e doenças: opção ou necessidade? EPAMIG, Belo Horizonte
Viana JB, Querino RB, Carvalho LC, Lima PD (2021) Sequence analysis of the internal transcribed spacer 2 (ITS2) region of rDNA for identifying Trichogramma species and evaluating genetic diversity. Braz J Biol 81:928–933. https://doi.org/10.1590/1519-6984.232362
Vickerman DB, Hoddle MS, Triapitsyn S, Stouthamer R (2004) Species identity of geographically distinct populations of the glassy-winged sharpshooter parasitoid Gonatocerus ashmeadi: morphology, DNA sequences, and reproductive compatibility. Biol Control 31:338–345. https://doi.org/10.1016/j.biocontrol.2004.07.001
Vieira JM, Querino RB, Cônsoli FL, Zucchi RA (2015) An integrative taxonomic approach to characterize Trichogramma marandobai (Hymenoptera: Trichogrammatidae). Zootaxa 4021(3):447–458. https://doi.org/10.11646/zootaxa.4021.3.4
von Felten A (2017) LAMP based identification of quarantine insects at the airports of Zürich and Geneva. Workshop for inspectors on tools available for inspection. EPPO Sand Hutton, York, pp 13–14
Wen J, Harris AJ, Ickert-Bond SM, Dikow R, Wurdack K, Zimmer EA (2017) Developing integrative systematics in the informatics and genomic era, and calling for a global Biodiversity Cyberbank. J Syst Evol 55:308–321. https://doi.org/10.1111/jse.12270
Wheeler QD (2004) Taxonomic triage and the poverty of phylogeny. Philos Trans R Soc B Biol Sci 359:571–583. https://doi.org/10.1098/rstb.2003.1452
Wheeler QD (2007) Invertebrate systematics or spineless taxonomy. Zootaxa 1668:11–18. https://doi.org/10.11646/zootaxa.1668.1.3
Wheeler QD, Valdecasas AG (2005) Ten challenges to transform taxonomy. Graellsia 61:151–160. https://doi.org/10.3989/graellsia.2005.v61.i2.17
Yara K, Sasawaki T, Kunimi Y (2010) Hybridization between introduced Torymus sinensis (Hymenoptera: Torymidae) and indigenous T. beneficus (late-spring strain), parasitoids of the Asian chestnut gall wasp Dryocosmus kuriphilus (Hymenoptera: Cynipidae). Biol Control 54:14–18. https://doi.org/10.1016/j.biocontrol.2010.03.006
Yu DSK, Achterberg CV, Horstmann K (2016) Taxapad 2016, Ichneumonoidea. Database on flash-drive. Nepean, Ottawa
Zamani A, Vahtera V, Sääksjärvi IE, Scherz MD (2021) The omission of critical data in the pursuit of ‘revolutionary’ methods to accelerate the description of species. Syst Entomol 46:1–4. https://doi.org/10.1111/syen.12444
Zhang AB, Muster C, Liang HB, Zhu CD, Crozier R, Wan P, Feng J, Ward RD (2012) A fuzzy-set-theory-based approach to analyse species membership in DNA barcoding. Mol Ecol 21:1848–1863. https://doi.org/10.1111/j.1365-294X.2011.05235.x
Zhang YM, Ridenbaugh RD, Sharanowski BJ (2017) Integrative taxonomy improves understanding of native beneficial fauna: revision of the Nearctic Peristenus pallipes complex (Hymenoptera: Braconidae) and implications for release of exotic biocontrol agents. Syst Entomol 42:596–608. https://doi.org/10.1111/syen.12233
Zhang YM, Stigenberg J, Meyer JH, Sharanowski BJA (2018) Multilocus phylogeny of the parasitic wasps in the tribe Euphorini (Hymenoptera: Braconidae) with revised generic classifications. PeerJ 6:e4783. https://doi.org/10.7717/peerj.4783
Zhang YM, Williams JL, Lucky A (2019) Understanding UCEs: a comprehensive primer on using ultraconserved elements for arthropod phylogenomics. Insect Syst Divers 3(5):3. https://doi.org/10.1093/isd/ixz016
Zucchi RA (1988) New species of Trichogramma (Hym., Trichogrammatidae) associated with the sugar cane borer Diatraea saccharalis (F.) (Lep. Pyralidae) in Brazil. Colloq INRA 43:133–140
Zucchi RA (2002) A taxonomia e o controle biológico de pragas. In: Parra JRP, Botelho PSM, Corrêa-Ferreira BS, Bento JMS (eds) Controle biológico no Brasil: parasitóides e predadores. Editora Manole, São Paulo, pp 17–24
Zucchi RA (2021) Taxonomia e controle biológico: uma relação mútua. In: Parra JRP, Pinto AS, Nava DE, Oliveira RC, Diniz AJF (eds) Controle biológico no Brasil com parasitoides e predadores na agricultura brasileira. Fundação de Estudos Agrários Luiz de Queiroz, Piracicaba, pp 55–77
Zucchi RA, Querino RB, Monteiro RC (2010) Diversity and hosts of Trichogramma in the New World, with emphasis in South America. In: Cônsoli FL, Parra JRP, Zucchi RA (eds) Egg parasitoids in agroecosystems with emphasis on Trichogramma. Springer, New York, pp 219–236
Acknowledgements
We are grateful for the valuable information regarding Brazilian regulations provided by Izabela Matosinhos and Vitor Domingues. RA Zucchi is a fellow of the National Council for Scientific and Technological Development (CNPq).
Funding
This study was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior—Brasil (CAPES)—Finance Code 001. The authors also thank the Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, grant numbers 2017/50334–3 and 2018/18965–6) and the Instituto Nacional de Ciência e Tecnologia de Hymenoptera Parasitoides (Proc. 65562/2014–0), for the support to one of the authors (VAC), and Fundação de Apoio à Pesquisa do Distrito Federal (Proc. n.º 435/2022), also for the support for one of the authors (RBQ). RAZ is a CNPq fellow.
Author information
Authors and Affiliations
Contributions
All authors contributed to the conceptualization of the article. EMS drafted the first version of the manuscript, and RAZ, RBQ, and VAC critically revised the text. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Edited by Élison Fabrício Bezerra Lima
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Shimbori, E.M., Querino, R.B., Costa, V.A. et al. Taxonomy and Biological Control: New Challenges in an Old Relationship. Neotrop Entomol 52, 351–372 (2023). https://doi.org/10.1007/s13744-023-01025-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13744-023-01025-5